
Глава 8. ГЕНЕТИКА ПОВЕДЕНИЯ
О возможности передачи особенностей поведения от родителей к потомкам было известно давно, поскольку при разведении сельскохозяйственных и домашних животных человек не только видел своеобразие их поведения, но и научился получать желаемые комбинации признаков поведения у новых пород. Наиболее отчетливые межпородные различия в поведении известны у собак.
Время существования научной дисциплины принято отсчитывать от появления первого исследования в этой области. Описанное в 1913 г. американской исследовательницей Адой Иеркс наследование комплекса злобности, пугливости и дикости у крыс (Rattus norvegicus) — первая экспериментальная работа по генетике поведения.
Началом самоопределения генетики поведения обычно считают год публикации первой обобщающей монографии американских ученых Дж.Фуллера и У.Томпсона "Генетика поведения" (Fuller, Thompson, 1960). Книга имела огромный успех у биологов разных специальностей. Оба автора по своему образованию не были генетиками, но, приступив к исследованию поведения, четко осознали значение генетического подхода. Не прибегая к сложным генетическим рассуждениям, они смогли убедительно показать необходимость оценки роли генотипа в формировании поведения и привести экспериментальные свидетельства этого. Еще одной работой, важной для становления этого направления, была монография Дж. Скотта и Дж. Фуллера по генетике поведения собак (см.: 8.6.1).
Значительную роль в формировании генетического подхода к анализу поведения сыграли работы сотрудников специальной генетической лаборатории в США, так называемой Джексоновской лаборатории в штате Мэн. Это учреждение — всемирно известный центр, основанный в 1929 г. генетиком К. Литтлом. В нем поддерживаются инбредные и селектированные линии мышей, число которых в настоящее время очень велико (см. также: 8.2.4). В этой коллекции имеется много десятков мутаций, затрагивающих строение мозга и поведение.
Джексоновская лаборатория может предоставить любое число животных, имеющих нужный исследователям генотип. Такая возможность позволила ученым разных стран не только скринировать множество линий и выявить межлинейные различия поведения и нейрохимических признаков, но и послужила основой для разработки новых подходов к изучению генетики количественных признаков (метода рекомбинантных инбредных линий — см.: 8.6.7.2 и метода картирования QTL — quantitative trait loci;, см.: 8.6.7.3), которые используются и в генетике поведения.
В нашей стране первым генетическим исследованием признаков поведения была выполненная в 20-х годах работа М.П.Садовниковой-Кольцовой. В ней была сделана попытка селекции крыс на скорость побежки в экспериментальной установке (см.: Крушинский, 1991). В последние годы жизни И.П.Павлова в Колтушах была организована лаборатория генетики высшей нервной деятельности, задачей которой должно было стать изучение генетических основ индивидуальных особенностей условнорефлекторной деятельности (т.е. типов ВНД) собак.
В этой лаборатории в конце 30-х годов Л.В.Крушинский начал исследования генетики поведения собак. По своему содержанию и методологии они практически не имеют себе равных и по сей день (см.: 8.6.1).
В нашей стране генетические исследования поведения животных проводились в нескольких лабораториях, в каждой из которых разрабатывалось свое оригинальное направление. Лаборатории, созданные в 30—40-е годы М.Е. Лобашевым (1907—1971) и В.К. Федоровым (1914—1972) в Колтушах (Институт физиологии им. Павлова АН СССР), изучали проблемы генетической детерминированности свойств нервной системы, а также (совместно с кафедрой генетики ЛГУ) вопросы сравнительной генетики поведения.
Следует также упомянуть об оригинальных генетических исследованиях Р.А.Мазинг, изучавшей проявление некоторых морфологических мутаций дрозофилы (Bar, eyeless, white и др.). Поведение мух, а именно их реакция на свет и выбор места для откладки яиц, при этих мутациях зависели от генетического фона линии, несущей мутации.
В Институте цитологии и генетики СО АН СССР (Новосибирск) под руководством Д.К.Беляева (1917—1987) в 60-е годы была начата селекционная работа по созданию доместицированной линии серебристо-черных лисиц. Отбор на спокойное и на агрессивное отношение к человеку вызвал к жизни сложнейшие перестройки эндокринной и нейромедиаторной систем организма, которые широко исследуются в настоящее время (Трут, 1987).
На биологическом факультете МГУ в лаборатории, созданной и возглавленной Л.В.Крушинским (в 1968 г. она получила название лаборатории физиологии и генетики поведения), была выведена линия крыс с высокой чувствительностью к звуку (линия Крушинского-Молодкиной — КМ) и исследована генетическая детерминированность аудиогенной эпилепсии (см.: 8.6.5.1) у этих животных. В этой же лаборатории под руководством Л.В.Крушинского были проведены исследования роли генотипа в формировании сложного адаптивного поведения — способности животных к экстраполяции направления движения стимула (см.: Крушинский, 1986).
Генетическое исследование поведения позволяет выяснить, в какой степени изменчивость интересующего нас признака поведения связана с изменчивостью генотипов данной группы животных, а в какой — с внешними по отношению к генотипу событиями, воздействующими на ЦНС, и следовательно, на поведение. Для этого необходимо использование так называемых генетических моделей — групп животных, состоящих из генетически идентичных (или почти идентичных) особей, имеющих определенные физиологические или биохимические особенности. С этой целью используют линии животных. Это могут быть инбредные линии (мышей и крыс), между которыми обнаружены межлинейные различия по поведению. Наряду с ними используются линии, сформированные путем селекции (искусственного отбора) на высокие и низкие значения какого-либо признака поведения.
Выявление межлинейных различий или выведение специальных линий — обычно первый этап исследования. Следующим шагом в классических исследованиях по генетике поведения бывает проведение скрещиваний животных из линий, обнаруживших контрастные значения признака.
Тестирование поведения гибридов первого поколения дает информацию о доминантном, промежуточном или рецессивном наследовании интересующего нас признака. Если данный признак определяется одним, двумя или тремя генами, то это можно определить по картине его распределения у гибридов второго поколения и потомков возвратного скрещивания. Если же в определении признака участвует большее число генов, то необходимо применять методы генетики количественных признаков (см.: 8.6.6).
Современный этап развития науки обогатил генетику поведения новыми методами. Кроме упомянутых выше методов рекомбинантных инбредных линий и QTL, это создание и исследование мозаичных и химерных животных, а также трансгенных организмов (см.: 8.5.3; 8.6.4.4). Очевидно, что подобные исследования можно проводить на объектах, которые хорошо изучены генетически.
Генетика поведения и нейрогенетика как ее часть сформировались в большой степени благодаря использованию плодовой мушки дрозофилы в качестве объекта исследования. Это относится и к исследованию процесса нейрогенеза, и к выявлению универсальных генных комплексов, работающих в живых системах разного уровня организации (дрожжевые грибки, амфибии, млекопитающие), названных гомеобоксом. Другой важнейшей группой работ, выполненных на дрозофиле, является исследование белка CREB, связанного с процессом формирования памяти и, по-видимому, имеющего универсальную природу (см.: 8.4.3.6).
Чрезвычайно перспективным экспериментальным объектом нейрогенетики и генетики поведения считаются мыши. Помимо задач общебиологического плана, связанных с исследованием генетических вариаций в строении мозга (см.: раздел 8.6.7) и их адаптивной значимости, на мышах широко изучаются нейробиологические основы процесса обучения. Исследование генетики поведения мышей дает возможность моделировать целый ряд неврологических и психических заболеваний человека, таких как эпилепсия, алкоголизм, депрессивные состояния, болезнь Альцгеймера и др. (см.: 8.6.5).
Основная задача генетики поведения — выяснение роли генетических факторов в определении особенностей поведения. Очевидно, что она состоит из нескольких достаточно самостоятельных проблем. Одна из них — определение относительной роли и взаимодействия генетических и средовых влияний при формировании поведения в онтогенезе. Вторая задача — исследование механизмов действия генов, определяющих формирование нервной системы. Третья задача связана с изучением механизмов реализации действия мутантных генов, затрагивающих функцию ЦНС, которые могут служить моделями заболеваний нервной системы человека. Четвертая задача — изучение генетико-популяционных механизмов формирования поведения и его изменений в процессе микроэволюции. Вторую и третью задачи нередко выделяют в направление, получившее название нейрогенетики.
Интеграция целостного, "организменного" и молекулярно-биологического подходов для создания возможно более полной картины роли генотипа в формировании мозга, в развитии его отдельных реакций и поведения в целом составляет общую задачу генетики поведения. Таким образом, две ее основные части — собственно генетика поведения и нейрогенетика дополняют друг друга, причем следует отметить, что в последнее время нейрогенетика начинает приобретать все большее значение.
Наряду с конкретными экспериментальными исследованиями, проведенными в середине и даже в начале века, истоком современной генетики поведения служит также феногенетика — направление, зародившееся в начале в 30-х годов (см.: 8.2.1). Задачей феногенетики является изучение закономерности становления признака под влиянием данного набора генов и определенной констелляции средовых воздействий (Астауров, 1968).
В настоящее время генетические исследования поведения и нейрофизиологических процессов проводятся по нескольким направлениям.
Условно они подразделяются на две группы: исследование функции гена на молекулярном и физиологическом уровне с последующим анализом влияния этого гена на поведение (подход "от гена к поведению") и изучение генетической компоненты изменчивости целостного поведения с последующим более детальным анализом феноменологии на уровне отдельных хромосом и генных комплексов или же отдельных генов (подход "от поведения к гену").
Исследования первой группы проводятся по ряду направлений. Во-первых, изучаются особенности влияния генов, кодирующих белки-предшественники пептидов, ферменты, структурные белки, определяющие общие и специфические признаки нервных и глиальных клеток (рецепторные структуры и молекулы, медиаторные системы, формирование клеточных органелл и отростков — дендритов и аксонов, синапсов и др.), а также белки, связанные с функцией ЦНС как целого. Сюда относятся также исследования влияния отдельных локусов, детерминирующих взаимодействие мозга и эндокринной системы, а также генов, участвующих в синтезе веществ химической сигнализации (например, гормона откладки яиц у аплизии, феромонов), и генов, детерминирующих специфику поведения у беспозвоночных животных, в особенности у насекомых (например, исследование мутаций, влияющих на половое поведение дрозофилы — см.: 8.4).
Во-вторых, это четкие и часто хорошо изученные поведенческие эффекты мутаций отдельных генов у млекопитающих. Это и мутации, мало влияющие на общую приспособленность организма в условиях содержания животных в лаборатории (их можно было бы отнести к "нейтральным"), и "вредные" неврологические мутации. Последние представляют большой практический интерес как модели генетических заболеваний нервной системы человека, а также как объект для изучения нормальных процессов развития и функции мозга (см.: 8.6.4).
Поиск четких "единиц" физиологической регуляции поведения заставляет исследователей переходить ко все более простым объектам.
Сложность генетической регуляции "примитивных" реакций у низших организмов будет показана в разделе 8.3, посвященном генетике "поведения" Escherichia coli и примитивных реакций нематоды Caenorhabditis elegans и др.
Вторая группа исследований использует подход "от поведения к генам", который нельзя назвать альтернативным предыдущему (см.: 8.6). Однако экспериментальные методы, используемые в таких работах, часто совсем иные, нежели при анализе работы отдельных генов. Именно в исследованиях такого рода, которые можно назвать генетикой поведения ''per se'' чрезвычайно важны выбор адекватного признака для анализа (см.: 8.2) и владение набором четких правил, составляющих основу генетического анализа поведения.
В 30-е годы на основе работ Б.Л.Астаурова, П.Ф.Рокицкого, Н.В.Тимофеева-Ресовского и др. сформировалась так называемая феногенетика — направление, анализирующее пути и правила становления признака в процессе развития. Если ранее наибольшее распространение имела лишь гипотеза "один ген — один фермент", то развитие феногенетики дополнило эти представления. Были сформулированы новые положения, каждое из которых имеет солидное экспериментальное обоснование.
В этот период были установлены два общих принципа отношений между генами и признаками: 1) каждый ген влияет на все признаки в организме, хотя его влияние на некоторые из них может быть исчезающе мало; 2) любой признак организма зависит от всех генов фенотипа в целом, даже если зависимость от некоторых незаметна. Иными словами, развитие каждого признака представляет собой цепь последовательных генных взаимодействий, проявляющихся в определенных условиях среды.
Как говорилось в разделе 8.1.2, одна из задач нейрогенетики (которая представляется в то же время задачей общей биологии развития) — исследование закономерности действия генов при дифференцировке клеточных элементов ЦНС и при формировании структур мозга в целом. В ходе развития организма формируются нейрохимические особенности разных отделов мозга и медиаторная специфичность нервных клеток, а также специализированные связи между нейронами. Очевидно, что детальное описание этих событий выходит за рамки данной книги. В разделах 8.2.10 и 8.5 дан краткий очерк некоторых представлений, существующих в этой области.
Особенности проявления признаков поведения, связанные со сложностью и многоступенчатостью процессов, лежащих в их основе, а также со сложностью и разнообразием влияний внешней среды на их формирование и проявление, представлены здесь кратко. Основными источниками более подробной информации по этим вопросам могут служить книги Эрман и Парсонса (1985), Трут (1978), руководство "Физиологическая генетика и генетика поведения" (Крушинский, 1981).
Влияние генов на фенотипические признаки может быть и непосредственным, и достаточно "далеким", опосредованным. Некоторые признаки, например первичная структура белков, есть отражение последовательности нуклеотидов в данном гене. Другие, более сильно отдаленные от первичного эффекта гена признаки, как правило, находятся под влиянием значительного числа генов. Так называемые количественные признаки — рост, вес, плодовитость и т.п., а также многие признаки поведения, связанные с функцией разных отделов мозга, также связаны с функцией конкретных генов непрямым образом.
Когда признак отдален от первичного эффекта гена несколькими "ярусами" биохимических процессов, то обнаруживается влияние некоего гена не только на исследуемый признак, но и на многие другие стороны строения, функций и поведенческих реакций организма. Это явление называется плейотропией. Описано множество генов со сложными плейотропными эффектами.
Классическим примером плейотропии, который способствовал накоплению экспериментальных свидетельств влияния генотипа на поведение дрозофилы, было впервые обнаруженное А.Стертевантом (1915) и подробно описанное в 1956 г. М.Бэсток влияние мутации yellow на уровень половой активности мух (см.: 8.4.3.2).
Широко известны многочисленные плейотропные эффекты мутации альбинизма, обнаруживающиеся не только у лабораторных мышей и крыс, но и у многих других видов животных и даже человека (см.: 8.6.4.1) — это нарушение остроты зрения, специфические особенности морфологии зрительных путей {полный перекрест волокон зрительного нерва).
Исследование мозга мышей с мутацией rd (дегенерация сетчатки), проведенное американскими исследователями Ц. и Р.Ваймерами, выявило ее плейотропный эффект. Дегенерация светочувствительных элементов сетчатки — палочек — определяется геном, расположенным на 5-й хромосоме. Процесс начинается с 20-го дня жизни. Специальными экспериментами на мышах-химерах (см.: 8.5.3.2) было установлено, что ген rd действует на уровне нервных элементов сетчатки, а не на уровне клеток пигментного эпителия. Анализ структур ЦНС выявил, что размер зубчатой фасции гиппокампа, а также количества гранулярных и корзинчатых клеток в этом отделе мозга у мышей с генотипом rd/rd были значительно ниже нормы. Механизм возникновения подобной аномалии пока неизвестен.
Описаны эффекты мутации гена brindled (Mo br), которые поначалу поставили исследователей в тупик. Симтомокомплекс этой мутации заключается в аномальном снижении двигательной активности и треморе, в светлой окраске шерсти и наличии у таких животных закрученных вибрисс. Оказалось, что снижение активности таких ферментов, как дофамин-бета-гидроксилаза (ответственное за снижение локомоции), тирозиназа в меланоцитах (светлая шерсть) и лизилоксидаза в волосяных фолликулах (аномальная структура вибрисс) вызвано общим фактором — ослаблением усвоения меди в кишечнике. Это, видимо, и является первичным эффектом данной мутации. Считается, что она в какой-то степени сходна с мутацией при так называемом синдроме Менкеса у человека (см.: Hay, 1985).
К плейотропным эффектам следует отнести также множественные отклонения от нормы в развитии мозга мышей при неврологических мутациях, например при мутации reeler, когда измененное в результате мутации развитие волокон радиальной глии влечет за собой нарушение миграции больших групп нейробластов (см.: 8.5.4).
Типичный плейотропный эффект — это последствия фенилкетонурии у человека. Фенилкетонурия характеризуется тем, что в организме отсутствует или малоактивен фермент фенилаланингидроксилаза, превращающий фенилаланин в тирозин. Если имеющийся недостаток тирозина возмещать богатой тирозином пищей, этот дефект не будет иметь последствий для жизнедеятельности в целом. Однако при этом в крови больных фенилкетонурией уровень фенилаланина оказывается повышенным, а продукты его обмена попадают в разные органы и ткани, в том числе и в мозг, нарушая их развитие. Вторичное влияние мутантного гена у человека проявляется в задержке умственного развития и особенностях темперамента, а также в изменении пигментации волос. Считается, что эти эффекты определяются участием ферментов, синтезирующих нейромедиаторы и меланин — пигмент волосяных фолликул.
Драматическими примерами сложных плейотропных влияний одиночных генов могут служить многочисленные мутации человека. Например, синдром Леш-Нихана связан с дефектом гена, ответственного за синтез гипоксантингуанинфосфорибозилтрансферазы, что вызывает ряд тяжелых расстройств — от подагры и заболевания почек до аномального поведения. Дети, пораженные этим заболеванием, обладают сниженным интеллектом и склонны к "самоистязанию", повреждая себе (часто необратимо) губы и пальцы. Характерно, что они испытывают при этом страдания, поскольку болевая чувствительность у них не изменена (см.: Эрман, Парсонс, 1984; Фогель, Мотульский, 1990)
В целом можно утверждать, что термин "плейотропия" относится к анализу эффектов генов на уровне организма и представляется сугубо описательным. Детальное исследование эффектов мутантных генов на уровне клетки, ткани и органа должно привести к выявлению конкретных путей нарушения физиологических функций, т.е. к расшифровке сущности плейотропных эффектов.
У генетики поведения были, особенно в начальный период существования, трудности, связанные со сложностью признаков поведения, которые продолжают существовать и сейчас. Несомненно, что для успеха их генетического исследования необходимо точно сформулировать задачу работы и выбрать признак, который представляет собой естественную "единицу" той или иной формы поведения. Очевидно, что успешно отыскать такой признак можно только на основе хорошего знания физиологии, нейрофизиологии и поведения.
Следует отметить, что логика построения физиологического исследования всегда требовала отыскания "типичного" варианта того или иного феномена. Очевидно, что для генетических исследований такой подход неприемлем. Суть генетического подхода состоит в оценке размаха изменчивости признака у данного вида, популяции или группы особей и в анализе происхождения этой изменчивости. Таким образом, столь необходимое для прогресса генетики поведения участие в таких работах физиологов до недавнего времени сдерживалось из-за существования подобного "типологического" подхода к предмету. В настоящее время данная проблема уже не стоит столь остро (см.: 6.4).
В период накопления фактов в генетике поведения внимание исследователей привлекали и разные виды животных, и разные признаки: предрасположенность к судорогам, общая возбудимость, локомоторная активность, ориентировочно-исследовательские реакции, разные аспекты репродуктивного поведения, классические и инструментальные условные реакции, реактивность к фармакологическим веществам.
Опыт, накопленный в первый период развития генетики поведения, можно суммировать следующим образом. Для исследования роли генотипа в формировании поведения следует выбирать такие признаки, которые легко поддаются количественному учету (например, четкие видоспецифические движения), либо такие, которые легко измерить по степени выраженности (например, уровень локомоторной активности, измеряемый по длине пройденного животным пути за фиксированное время опыта).
Многие признаки поведения сильно зависят также от ряда внешних по отношению к нервной системе факторов, например от сезона года и/или от гормонального фона. Это вызывает дополнительные трудности при проведении генетических исследований.
Основу и первый этап генетического исследования признаков поведения и/или физиологических признаков составляет оценка их генетической изменчивости. Очевидно, что если речь не идет о группе инбредных, т.е. генетически идентичных друг другу животных, то генетический компонент изменчивости признаков поведения всегда присутствует, составляя большую или меньшую долю фенотипической изменчивости.
Проблема определения объекта исследования в генетике поведения решается, как правило, выбором либо группы инбредных линий для сравнения, либо проведением селекционного эксперимента на высокие и низкие значения этого признака. Какую именно генетическую модель необходимо выбрать, определяется конкретными целями исследования и спецификой изучаемого фенотипического признака.
Признаки организма могут варьировать, обнаруживая фенотипическую изменчивость в пределах нормы реакции, размах которой определен генотипом. Генетическая изменчивость особей популяции обнаруживается по огромному числу признаков, в число которых входят и альтернативные (наличие — отсутствие признака), и так называемые количественные, когда величина признака определяется большим числом пар аллелей, вклад каждой из которых определить достаточно трудно. Использование методов рекомбинантных инбредных линий и QTL (см.: 8.6.6) дает по меньшей мере теоретическую возможность локализации целого ряда генов, суммарный эффект которых определяет данный количественный признак.
Помимо изменчивости в пределах нормы реакции, характерной для данного генотипа, а также помимо генетической изменчивости, связанной с гетерогенностью аллельного состава данной популяции, для признаков поведения характерна еще одна, специфическая форма изменчивости, которая не может быть прямо отнесена ни к первой, ни ко второй категории. Речь идет об изменчивости признаков поведения животных, связанной с воздействием индивидуального опыта, т.е. с разными формами сенситизации, привыкания, обучения, формирования представлений и т.д. Иными словами это то, что составляет специфику мозга как структуры, обеспечивающей научение, и что тоже является предметом внимания генетики поведения.
Напомним, что в соответствии с классическими представлениями этологии подобную пластичность можно обнаружить при выполнении животным поисковой фазы поведения, тогда как собственно инстинктивные акты поведения — фиксированные комплексы действий (ФКД, см.: 3.3), входящие в завершающий акт, пластичностью практически не обладают. Каждый из ФКД может служить достаточно дискретной и четкой "единицей" для анализа закономерностей его генетического контроля (см. также: 8.2.5).
Селектированные и инбредные линии представляют собой основной материал для работы в области генетики поведения. Селекция мышей и крыс на степень выраженности ряда признаков поведения была успешной. В настоящее время осуществлены селекционные программы, которые охватили практически весь диапазон признаков поведения, показавших генетическую изменчивость, — от скорости проведения нервного возбуждения в нервно-мышечном синапсе (см.: Лопатина, Пономаренко, 1987) до высокой и низкой способности к обучению (см.: 8.6). В то же время с помощью селекции формируются линии, которые состоят из животных, сходных по выраженности исследуемого признака и иногда ряда других признаков, но не обладающих генетической однородностью. Это затрудняет использование их в целях идентификации генов, влияющих на данный признак.
В ряде случаев удается провести работу по выведению инбредных селектированных линий, т.е. полностью гомозиготных животных с контрастными в случае двух линий величинами интересующего нас признака. Инбридинг (близкородственное скрещивание) начинают после того, как получены неперекрывающиеся межлинейные различия по интересующему нас признаку. Таких линий немного. В качестве примера можно привести инбредные линии мышей Short- и Long sleep, у которых после гипногенной дозы этанола развивается либо короткий, либо длинный период сна. Основную же массу составляют инбредные линии мышей и крыс, выведенные ранее, до начала активных исследований в области генетики поведения. Такие линии были необходимы для исследований по иммунологии, гистосовместимости, онкологии и др, (см, также: Бландова и др., 1983).
Инбредные линии мышей и крыс получаются при размножении животных путем братско-сестринского скрещивания в течение примерно двадцати поколений, Полезные свойства этих линий позволяют использовать их для генетического анализа и выявления локусов, влияющих на поведение. Образно говоря, инбредная линия представляет собой неумирающий клон, состоящий из генетически идентичных друг другу особей.
Ниже перечислены основные свойства, отличающие инбредные линии от генетически гетерогенных аутбредных (Wimer, 1992).
1. Изогенность. Все животные инбредной линии генетически идентичны друг другу практически по всем локусам. Это означает, что для получения генетического "профиля" линии соответствующее определение можно сделать на одном животном. Для начала дочерней колонии инбредной линии можно использовать только одну пару.
2. Гомозиготность. Инбредные линии гомозиготны практически по всем локусам. У них нет никаких "скрытых" генов, которые не выявлялись бы при размножении линии "в себе". Естественно, что никаких изменений генетического состава, связанных с дрейфом генов или отбором, в инбредной линии произойти не может.
3. Фенотипическое подобие. Поскольку инбредная линия не может иметь генетических отклонений, фенотипическое единообразие таких животных по многим признакам также оказывается более высоким, чем в аутбредных стоках. Это относится в первую очередь к признакам с моногенным или олигогенным (т.е. когда задействовано малое число генов) определением. Так, например, после введения гексобарбитала мышам линии Balb/c их сон продолжается в среднем 41±2 мин, а у животных аутбредного стока Swiss примерно столько же — 43±15 мин, но разброс величин у последних был значительно большим. Если предполагается оценить изменение длительности сна в ответ на какое-либо воздействие, то, согласно простым расчетам, для получения достоверного результата достаточно использовать лишь 15 мышей линии BALB/c, но 290 аутбредных мышей Swiss.
Инбредные линии могут иметь повышенную чувствительность к средовым воздействиям и, следовательно, фенотипическое сходство особей по признакам поведения, имеющим полигенную природу, может нарушаться (см. п. 7).
4. "Индивидуальность" линии. Каждая инбредная линия генетически уникальна; ее поведенческий и неповеденческий фенотипы всегда будут непохожи на фенотипы любой другой линии. Это означает, что практически для всех проявлений поведения можно обнаружить межлинейные различия. Из этого следует очень важный вывод о том, что выявление различий между линиями по поведению требует в дальнейшем исследования их физиологических и биохимических причин.
5. Долговременная стабильность. Инбредные линии остаются генетически стабильными в течение больших отрезков времени. Примером может служить постоянство генетического состава двух родственных сублиний — CS7BL/6 и C57BL/10, которые, хотя и ведутся независимо уже более 50 лет, но остаются, как и в начале, на 97% генетически сходными.
Нарушение генетической стабильности инбредной линии может произойти в результате одной из трех причин — наличия остаточной гетерозиготности, новых мутаций и генетического "заражения". Остаточная гетерозиготность есть следствие того, что инбридинг даже в течение многих поколений по какой-то причине не может привести к полной гомозиготности. Возникновение мутаций — событие достаточно редкое, но есть случаи, когда появлялись мутации, влияющие и на поведение.
6. Возможность идентификации. Поскольку каждая линия имеет свой генетический "профиль", не составляет большого труда проверить принадлежность животного к этой линии. Это особенно важно при подозрении на засорение линии посторонним генетическим материалом. Подобное "заражение" линии может, к сожалению, случаться при ошибках или небрежности в разведении.
7. Чувствительность. Гомозиготность особей инбредной линии считается причиной того, что эти животные часто оказываются более чувствительными к внешним воздействиям. Инбредные животные гомозиготны в том числе и по аллелям, которые не обязательно способствуют повышению их жизнеспособности, а нередко, наоборот, ведут к ее снижению. Противоположное явление, а именно малую чувствительность гибридов первого поколения к средовым влияниям, объясняют наличием генетического буфера, т.е. формированием гетерозиготного организма, у которого каждый аллель, вызывающий снижение жизнеспособности, с большой вероятностью имеет "парой" аллель другого происхождения, который такого действия не оказывает.
Отсутствие подобных "буферных" свойств у гомозиготного генотипа может быть причиной слабой защищенности инбредного организма от внешних, нарушающих развитие влияний. Это же явление лежит в основе иногда обнаруживаемой повышенной чувствительности животных инбредных линий к средовым воздействиям, что может приводить к появлению фенотипических различий между инбредными животными одной линии, противодействуя тем самым свойству фенотипического подобия, о котором говорилось выше (п. 3).
8. Использование в разных странах. Поскольку инбредные линии обладают свойствами изогенности, гомозиготности и возможностью быть идентифицированными, их можно использовать в лабораториях разных стран без опасения каких-либо генетических изменений в них, связанных с малым числом основателей колонии (как это имеет место при работе с аутбредными стоками, например селектированными линиями).
9. Использование данных прошлых лет. Данные по инбредным линиям в силу их генетической стабильности можно сравнивать с результатами, полученными на тех же линиях, но ранее. Это облегчает планирование и проведение новых исследований.
Сочетание слов "изменчивость ФКД" находится в видимом противоречии с тем, что уже говорилось выше о стабильном характере простейших элементов инстинктивного поведения. Как разрешается такое противоречие? Несмотря на то что в целях описания и анализа поведение удобно рассматривать как состоящее из отдельных "форм" и/или актов, в реальности оно представляет собой непрерывный процесс, "поток", течение которого обозначается определенными конкретными актами.
Из этологического анализа, а также из данных генетико-популяционных исследований инстинктивного поведения следует, что природная генетическая изменчивость затрагивает не "рисунок", или паттерн (pattern), ФКД, а преимущественно частоту его выполнения и пороги их активации, т.е. их пространственно-временные характеристики. Первым внимание к этой проблеме привлек А.Меннинг (см.: Manning, 1967).
Таким образом, частота выполнения поведенческих актов (большая часть из которых — типичные ФКД), а также пороги их провокации в тех или иных условиях характеризуются высоким уровнем фенотипической и генотипической изменчивости поведения.
Иллюстрацией этого положения может служить изменчивость ФКД, входящих в репертуар поведения ухаживания самца дрозофилы (рис. 8.1).
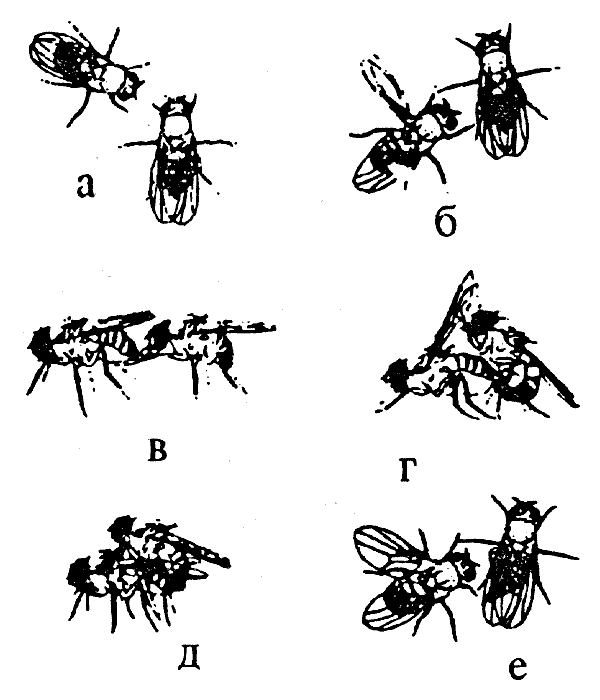
Рис.8.1. Стадии ритуала ухаживания у дрозофилы (с черным кончиком брюшка — самец): а — ориентация; б — отставление крыла и вибрация; в — "облизывание" гениталий; г — садка; д — копуляция; е — невосприимчивая самка отталкивает самца (оттягивает яйцеклад, поворачивая конец брюшка вбок) Все стадии обнаруживают межвидовую и внутривидовую изменчивость по продолжительности, а стадия вибрации характеризуется и видоспецифическим "рисунком" брачной песни
Этот ритуал условно подразделяют на 4 стадии: ориентация, вибрация, лизание и копуляция. При мутациях может изменяться как длительность, так и интенсивность данного ФКД. Межвидовые различия в поведении близкородственных видов, как правило, также связаны с подобной изменчивостью поведения.
У млекопитающих (мышей, крыс, собак и др. видов) значительную часть межлинейных и межвидовых различий в поведении можно свести к различиям в частоте выполнения видоспецифических ФКД. Это особенно четко проявляется в поведении линейных лабораторных грызунов, селектированных на высокие и низкие показатели признаков поведения. В качестве примеров можно привести различия в интенсивности проявления материнского поведения — подтаскивание детенышей к гнезду (Hurniket al., 1973), число вертикальных стоек (исследовательское поведение) (van Abeelen, 1974), проявления агрессивности (например, вращение хвостом). Множество примеров межлинейных различий содержится в любом обзоре по генетике поведения (Меннинг, 1982; Эрман, Парсонс, 1984). У межвидовых гибридов, полученных от видов, в естественных условиях не скрещивающихся, проявления ФКД имеют ряд особенностей.
Одно из первых подробных исследований гибридных форм провел К.Лоренц. У межвидовых гибридов уток выявляются такие ФКД, которых практически не бывает в репертуаре обеих родительских форм. Можно полагать, что генотип данного вида позволяет обеспечить физиологические механизмы таких стереотипных действий, но в силу каких-то причин они не выявляются фенотипически.
В 1975 г. внимание к этой проблеме привлек П.Лейхаузен (Leyhausen, 1975) при исследовании закономерностей проявления некоторых видоспецифических особенностей типичного ФКД кошачьих — "смертельного укуса" (см.: 7.5.3). Конкретной причиной, почему не все ФКД, на которые "способна" ЦНС данного вида, реализуются в репертуаре его поведения, могут быть различия в физиологических порогах их активации. В пользу этого предположения говорят исследования А.Хааса, посвященные поведению шмелей, о которых рассказывает в своей книге Лейхаузен.
Хаас наблюдал, как эти насекомые отыскивают свои гнезда. Изменяя расположение гнезд, он вызывал у шмелей состояние повышенного возбуждения и "стресса". Их поведение включало стереотипные элементы, свойственные другим видам шмелей того же рода. Такая потенциальная готовность к выполнению ФКД (в норме не характерных для вида), позволила предположить, что каждый вид изначально обладает всем "родовым" репертуаром поведенческих актов, но в обычных ситуациях реализует только определенную его часть. Экстренные ситуации, приводящие к высокому уровню активации ЦНС, могут выявить и остальные элементы "родового стереотипа".
Дезорганизацию гнездостроительного поведения наблюдал У.Дилгер (см.: Эрман и Парсонс, 1985) в эксперименте с гибридными особями попугаев-неразлучников. Они были получены от скрещивания таких двух видов, которые при постройке гнезда совершали различные действия: один из видов переносил гнездовой материал (в данном случае полоски бумаги) в клюве, другой — между перьями хвоста. Иными словами, два скрещивавшихся вида различались по ФКД при осуществлении врожденного поведенческого акта.
Гибридные особи в первый сезон размножения оказались не в состоянии построить гнездо, поскольку не могли справиться до конца с фиксацией гнездового материала. Они брали полоски бумаги в клюв (как один из родителей), затем пытались засунуть их между перьями, но делали это не так умело, как второй родитель. В результате постройка гнезда у этих птиц стала налаживаться только в последующие сезоны размножения, но их действия оставались нечеткими, и гнезда получались плохие.

Рис.8.2. Некоординированные действия неразлучника-гибрида по фиксации гнездового материала
Подобный феномен описан и в брачном поведении межвидовых гибридов рыбок-меченосцев. Самцы-гибриды первого поколения от сжрещивания Xiphophorus helleri и X.montezumae cortezi в состоянии высокого полового возбуждения демонстрируют последовательность действий, характерную для самцов X.helleri, тогда как при меньшем уровне возбуждения их поведение похоже на X.montezumae.
Все описанные и сходные с ними наблюдения (например, рисунок песни у гибридных сверчков — рис. 8.3) остаются пока вне поля зрения нейрогенетиков и нейроэтологов, однако можно полагать, что большинство их следует объяснять физиологической модуляцией порогов провокации видоспецифических ФКД.
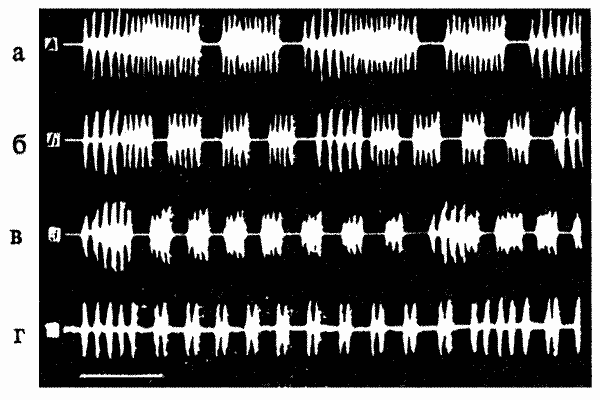
Рис.8.3. Рисунок песни у сверчков разных групп: а — Т.oceanicus, г — Т.commodus и двух групп их реципрокных гибридов — б и в
А.Меннинг еще в 1967 г. (Manning, 1967), анализируя генетические различия в половом поведении дрозофилы, высказал предположение, что генетическая изменчивость порогов и частоты выполнения ФКД может быть основой процесса микроэволюционных изменений поведения.
В таком случае возможно, что одним из механизмов формирования индивидуальных различий в поведении, связанных с генотипическими различиями, является разная "легкость" провокации тех или иных ФКД.
Изменчивость порогов проявления видоспецифических движений может зависеть от уровня возбудимости отдельных структур мозга и/или его отдельных медиаторных систем. Модуляция порогов ФКД, по всей видимости, происходит и при действии фармакологических веществ, изменяющих поведение животных. Это дает основание надеяться, что анализ роли генотипа в эффектах фармакологических препаратов на уровне целого организма перспективно рассмотреть еще и с этой точки зрения. Возможно, что микроэволюционные изменения поведения, описанные в многочисленных наблюдениях (Панов, 1978), действительно реализуются на основе динамических изменений в порогах проявления ФКД.
В работе Л.В. Крушинского (подробнее см.: 8.6.1) по наследованию оборонительных реакций у собак впервые было показано, что уровень возбудимости ЦНС влияет на экспрессивность и пенетрантность отдельных поведенческих реакций (Крушинский, 1991).
Экспрессивностью называется степень фенотипического проявления эффектов аллеля определенного гена у разных генетически сходных между собой особой» В основу различий экспрессивности могут лежать многие причины, в частности влияние средовых факторов или генетического фона. Если экспрессивность очень изменчива (вплоть до отсутствия проявления признака у отдельных особей), вводится дополнительная характеристика проявления действия гена — пенетрантность (частота проявления аллеля определенного гена у разных особей группы родственных организмов).
Примером изменчивости признаков поведения, которые связаны с экспрессивностью и пенетрантностью, могут быть количественные вариации в проявлении ФКД исследовательской активности, пищевого поведения, в интенсивности репродуктивного поведения, которые можно наблюдать у животных идентичных генотипов, например у мышей и крыс инбредных линий.
Внутрилинейная изменчивость признаков поведения определяется очень многими факторами. Один из них — это влияние соседства сородичей в жилой клетке, что редко учитывается исследователями. Как известно, лабораторные мыши и крысы относятся к видам, живущим в сообществах. Изоляция такого животного (в особенности самца) вызывает мощные изменения нейрохимических процессов в организме, а также изменения в поведении. Содержание животных в группе выявляет различия, определяющиеся положением особи во внутригрупповой иерархии. Это лишь один из примеров проявления изменчивости поведения особей с идентичными генотипами. Причины этого явления необходимо знать для оценки роли генотипа в формировании признаков поведения.
Еще один пример — различия в экспрессивности такого признака, как агрессивность. Американский исследователь Фом Сааль с сотрудниками показали, что к моменту рождения у эмбрионов мыши мужского пола уровень тестостерона в крови втрое выше, чем у самок, что связано со скачкообразным началом функционирования у них семенников. В проведенном эксперименте эмбрионов непосредственно перед родами извлекали методом кесарева сечения. При этом определялось относительное расположение в матке каждого из них и устанавливалось, кто были "соседями" данного плода (рис.8.4).
Напомним, что у грызунов двурогая матка, и эмбрионы в ней располагаются линейно. Выросшие взрослые самцы, соседями которых в матке также были самцы, оказались более агрессивными, чем те, которые в эмбриональном периоде располагались рядом с самками. В дальнейшем эти исследователи сравнили поведение и ряд морфологических характеристик самок, имевших соседями по матке двух самцов, с самками, соседками которых были две самки.
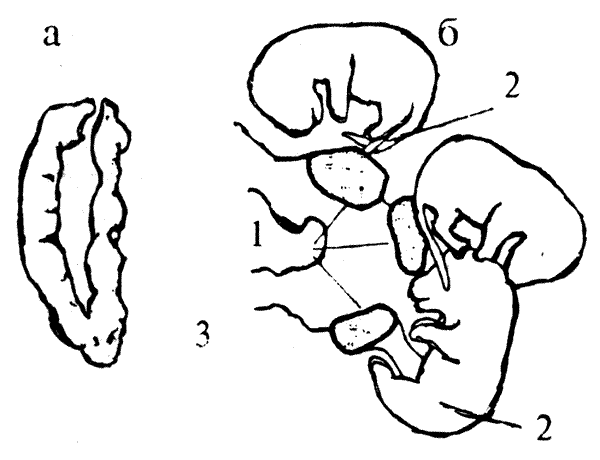
Puc.8.4. Расположение эмбрионов в двурогой матке грызуна: а — интактная матка; б — эмбрионы, извлеченные из нее; 1 — плацента; 2 — эмбрион, 3 — пуповина
У первых отмечались некоторые черты "маскулинизации" поведения и повышенный уровень тестостерона в крови. Несмотря на то, что у каждого из эмбрионов имеется собственная плацента, считается вполне вероятным влияние на данный плод состава крови "соседа" по матке. Именно этим, точнее повышенным пренатальным уровнем тестостерона в крови эмбрионов-самок и объясняют изменчивость уровня агрессивности у взрослых животных. Таким образом, это еще один пример того, что животные с одинаковым генотипом могут отличаться вариабельностью поведения, причина которой кроется исключительно в средовых, т.е. внешних по отношению к геному факторах.
Нередко оказывается, что в поведении и/или в неврологическом статусе организма-носителя мутации имеются многочисленные отклонения, которые могут наводить на мысль о существовании причинных связей между ними. Однако при выявлении нескольких эффектов данного мутантного гена зачастую высказывают мнение о существовании между ними причинно-следственной связи. Чтобы избежать при этом возможных ошибок, плейотропные эффекты мутации необходимо исследовать всесторонне.
Как уже говорилось выше (см.: 8.2.2), у лабораторной мыши при мутации brindled, локализованной в X-хромосоме, отмечаются "вьющиеся" вибриссы, "ослабленная" окраска шерсти, тремор, а также ряд других аномалий (Hay, 1982). Эти проявления мутантного гена являются следствием нарушений в усвоении меди — микроэлемента, необходимого для многих биохимических реакций. Введение в пищу новорожденным животным соединений меди практически компенсирует выявленные аномалии.
Существует несколько десятков форм наследственных заболеваний человека, вызванных нарушениями обмена, в число симптомов которых входят также неврологические и психические отклонения (см.: Эрман, Парсонс, 1984). Знание "метаболических" причин этих заболеваний позволяет более рационально подходить к поиску путей их коррекции.
Влияние внешних условий на изменчивость поведения исследуется почти исключительно на инбредных линиях животных, главным образом на мышах. Наряду с факторами внешней среды и генотипом у млекопитающих в формировании признака участвует еще один важнейший параметр — так называемый материнский эффект. Под этим термином понимают пре- и постнатальные влияния материнского организма, которые в свою очередь также зависят от генотипа. Эти влияния должны учитываться как еще один потенциальный источник изменчивости поведения взрослого животного.
Наиболее простые примеры влияния материнского организма на физиологические реакции и поведение потомства — это эффекты различных пренатальных воздействий на самку (стресс, фармакологические агенты), выраженность которых может зависеть от ее генотипа.
Экспериментально показано, что при соблюдении стандартных условий выращивания как матерей, так и их потомства, физиологические реакции и поведение последнего зависят от типа влияний "материнской среды". Эта влияния могут существенно сказываться не только на внутрилинейной, но и на межлинейной изменчивости. Иными словами, влияние иматеринской среды" инбредной линии А на формирование признака может быть иным, чем влияние на этот признак "материнской среды", свойственной генотипу B.
Влияние материнской среды складывается из 3 компонентов: цитоплазматического, внутриутробного и постнатального. Цитоплазматический компонент (или материнский эффект per se} обнаруживается не только у млекопитающих, но и у тех животных, развитие которых проходит вне материнского организма. Он заключается в том, что яйцеклетка и сперматозоид при слиянии привносят разные количества цитоплазмы, причем белки, синтезируемые на основе материнских мРНК, обнаруживаются в зародыше и после начала дробления. Кроме того, цитоплазма яйцеклетки передает зародышу значительно большее количество митохондриальной ДНК, чем цитоплазма спермия. Это означает, что цитоплазматические влияния могут иметь и генетическую природу.
Внутриутробный компонент материнского эффекта определяется генотипом матери, а также специфическими воздействиями, которые испытывает ее организм. Самки разного генотипа, даже выращенные и содержащиеся в одинаковых условиях, создают потомству разную "материнскую среду", что может повлечь за собой различия в фенотипе потомства.
Постнатальный компонент связан с тем, что детеныши млекопитающих, в особенности незрелорождающихся видов, к которым принадлежат мышь и крыса, длительное время зависят от материнской заботы, которая включает вылизывание потомства, поддержание чистоты и тепла в гнезде, количество и состав материнского молока, а также комплексное воздействие обеих групп факторов одновременно.
Разработаны экспериментальные приемы, с помощью которых можно оценить относительный вклад каждого из этих компонентов в фенотипическую изменчивость признака поведения. В основе столь развернутого методического подхода к проблеме лежат работы французской исследовательницы М. Карлье (Carlier et al., 1992).
Сравнивается выраженность исследуемого признака у следующих групп животных:
1) реципрокных гибридов первого поколения;
2) животных, воспитанных матерями разных генотипов (перекрестное вскармливание);
3) животных, выращенных из эмбрионов, пересаженных в матку самки либо своего, либо другого генотипа;
4) животных, которые развились из яйцеклеток, созревших в яичниках, пересаженных в организм самки другого генотипа.
Подобная экспериментальная процедура представляется громоздкой, формальной и кажется даже необязательной. В то же время знание возможных источников влияния на фенотип позволяет оперировать понятием "признак поведения" с большей уверенностью, а также оценивать разные компоненты изменчивости с большей точностью, чем при игнорировании таких эффектов. Хотя бы частичное применение перечисленных подходов, несомненно, важно в случаях, когда анализируют влияние генотипа на формирование мозга и поведения млекопитающих.
Успешность вьщепения "единиц" для анализа генетического контроля поведения требует выполнения следующих общих эмпирических правил исследования.
1. По возможности необходимо проводить повторные замеры признака с целью уменьшения ошибки, связанной с индивидуальной изменчивостью, разумеется, если такое повторение не влияет на выполнение теста.
2. Наблюдения должны быть длительными. Например, при оценке исследовательской активности в установке "открытое поле" автоматическая регистрация параметров поведения позволяет удлинить период наблюдения, увеличив тем самым точность снимаемых показателей.
3. Желательно одновременно регистрировать возможно большее число признаков поведения, что позволяет использовать современные методы оценки данных, выявляя неочевидные коррелятивные связи. Это часто способствует выявлению не только генетической, но и физиологической "архитектуры" признака.
4. Следует использовать автоматические системы регистрации поведения и унифицировать условия манипуляции с ними. Важно быть уверенным в отсутствии явных и скрытых инфекций у подопытных животных, для чего нужна их специальная подготовка к эксперименту, задача которой — устранение патогенных факторов.
5. При исследовании реакций животных на введение фармаколо-гаческих агентов рекомендуется использовать не одну, а ряд дозировок, что позволит более полно оценить чувствительность животных разных генотипов.
Что важнее для организма — генотип или воздействия среды? Эта проблема — соотношение врожденного и приобретенного в поведении — затрагивалась в главе 7. В настоящее время подобного вопроса практически никто не задает: сформулирована и "работает" современная эпигенетическая концепция (Рэфф, Кофмен, 1988; см. также: 8.5).
В оценке соотношения генотипа и среды для формирования поведения наиболее важным является определение степени лабильности или, наоборот, ригидности любого изучаемого признака поведения. В этом смысле и полемика "nature-nurture", и ранние эпигенетические представления (см.: Дьюсбери, 1981) имеют только исторический интерес. Напомним, что в главе 7 (см.: 7.2) мы дали краткое изложение того, как классическая этология решает проблему соотношения врожденного и приобретенного.
Как следует из современных представлений об относительной роли генотипа и средовых условий в формировании признака (см.: 8.5), поведение животного формируется при взаимодействии продуктов экспрессии генов (т.е. генотипа) со средой, которая в свою очередь порождается работой генов на предшествующей стадии и констелляцией внешних условий. Различный "удельный вес" вклада генотипа и среды в формирование признаков поведения зависит от типа генетической программы (см.: 8.5). Нужно также помнить, что поисковый компонент инстинктивного акта включает в себя значительное число пластичных, модифицируемых реакций, тогда как завершающий акт жестко детерминирован и лишь изредка обнаруживает изменчивость.
Разные уровни "пластичности" или "ригидности" одной и той же формы поведения иногда встречаются у животных близких видов. По-видимому, это может определяться особенностями их генотипов, точнее, предысторией микроэволюционного развития анализируемой формы поведения. Речь идет о том, какое давление отбора испытывал набор признаков данного вида или популяции в связи с экологическими особенностями конкретной среды обитания.
Примером, иллюстрирующим это положение, могут служить особенности пищевого поведения разных подвидов оленьей мыши (Peromyscus), описанные американским исследователем Дж. Кингом (King, 1977). Он анализировал пластичность пищевого поведения у двух подвидов P.maniculatus — северной (P.m.borealis) и южной (P.m.blandus) формы. Места обитания обеих форм характеризуются сухостью климата и большими колебаниями температуры, но в полупустыне, где живет P.m.blandus, растительность разнообразнее, и существует много животных, составляющих конкуренцию за пищу. Было показано, что P.m.blandus в значительной степени специализируется на питании определенными видами растений, а набор поведенческих актов, которые эти зверьки используют при кормлении, также ограничен. В то же время поведение P.m.borealis более пластично, что диктуется условиями обитания этой формы. Эксперименты показали, что процедура "хэндлинга" (взятия детеныша в руки), вызывающая умеренный неонатальный стресс, усиливает выявленные различия в поведении.
Последний пример иллюстрирует общее явление различия в реакциях на внешние воздействия у носителей разных генотипов.
Таким образом, поведение как объект исследования генетики представляется крайне сложным. Путь "от гена к признаку", который бывает достаточно короток у примитивных организмов, оказывается сложным и многоступенчатым при формировании нервной системы и поведения. Оценка этого пути у животных, организм которых состоит из одной клетки, а также у животных с относительно просто устроенной нервной системой, вносит некоторую упорядоченность в наши знания о генетическом контроле функций нервной системы.
В случае очень многих мутаций экспрессия аномального гена прерывает эмбриогенез, и зародыш гибнет. Однако в ряде случаев оказывается возможным реконструировать функцию нормального аллеля такого гена. Если удастся выделить мутации нескольких генов, экспрессирующихся на последовательных стадиях развития, то это может дать достаточно полную картину развития нервной системы. Подробная информация такого рода уже известна, она получена на интереснейшем генетическом объекте — микроскопической (2—3 мм) почвенной нематоде Caenorhabditis elegans (Кайданов, Лучникова, 1981; Rankin, Beck, 1992).
То, что удается узнать о генетическом контроле развития нервной системы и поведения, а также о генетическом контроле пластичности поведения у относительно простых объектов, делает, как мы надеемся, более понятной роль генотипа в формировании поведения насекомых и млекопитающих. Ниже приведены некоторые сведения по генетическому контролю физиологических реакций и "поведения" у бактерий, инфузорий и C.elegans.
Разумеется, можно лишь весьма условно говорить о наличии "поведения" у бактерии. Очевидно, что движение бактерии как ответная реакция на градиент химических веществ — аттрактантов и репеллентов — является простейшим поведенческим актом, контролируемым генетически. Если жгутики — "органы движения" бактерии — вращаются в направлении "против часовой стрелки", они собираются в единый пучок, благодаря чему она совершает плавное поступательное движение. Если же направление вращения жгутиков сменяется на "по часовой стрелке", то пучок рассыпается, бактерия "спотыкается", останавливается и меняет направление движения.
Наличие рецепторов химических веществ позволяет бактериальной клетке, этому одноклеточному организму, получать информацию об их концентрации. В положительном градиенте привлекающих или отрицательном — неприятных веществ она движется достаточно плавно, практически не останавливаясь.
Таким образом, в репертуар "поведения" клетки-организма входят: возможность узнавания стимула, способность к обработке информации о его параметрах, формирование ответной реакции. Был описан целый ряд мутаций Escherichia соli, при которых перечисленные выше реакции нарушались. Условно их подразделяют на следующие категории:
а) нарушение рецепции (т.е. восприятия сигналов от аттрактантов или репеллентов); вещества, сходные по строению (сахара, аминокислоты) действуют на свои специфические рецепторы, мутации которых независимы друг от друга;
б) нарушение передачи сигнала от одного или более специфических рецепторов к жгутикам;
в) мутации, при которых жгутики способны к движению только в направлении "по часовой стрелке", что ведет к "спотыканиям"; реакции на химические вещества сохраняются, но при значительно более высоких концентрациях, чем в норме;
г) мутации, при которых жгутики способны к движению только "против часовой стрелки", и клетка, следовательно, может двигаться только по прямой;
д) нарушение работы жгутиков или их способности к вращению.
Аналитические исследования, проведенные в 70—80-е годы, показали, что на этом объекте можно исследовать наиболее примитивные формы адаптации и фиксации предшествующего опыта, а также влияния условий окружающей среды на "поведение" клеток. Подробнее эти вопросы изложены Л.Эрман и П.Парсонсом (1984).
Простейшие, и в частности инфузории, — это перспективный объект для исследования генетики наиболее примитивных реакций. Эти организмы крупнее и неизмеримо сложнее бактерий. В качестве генетического объекта обычно использовались инфузории рода Paramecium, в частности один из наиболее крупных видов — P.aurelium.
Инфузории могут размножаться как бесполым (митотическим делением), так и половым путем. Половое размножение — конъюгация — обеспечивает генетическую изменчивость популяции, тогда как аутогамия — присущий простейшим способ самооплодотворения — позволяет получать полностью гомозиготных особей уже во втором поколении.
Движение инфузории осуществляется с помощью ресничек, покрывающих всю поверхность тела. Движение ресничек координируется общим физиологическим механизмом, связанным с возбудимой мембраной.
Мутации, выявленные у инфузории, влияют главным образом на характер движения. Особи "дикого типа" в норме перемещаются вперед короткими "проплывами", которые прерываются поворотами на 90o. Такие повороты рассматриваются как проявление спонтанных реакций избегания.
Генетические нарушения — мутации — выявляются именно в характере движений этих животных (рис. 8.5). При рецессивной мутации fast-2 инфузория очень быстро перемещается, совершая при этом большое число поворотов. Группа мутаций pawn (3 локуса, 62 мутации) характеризуется полным отсутствием реакции избегания, и клетка может двигаться только вперед (как пешка в шахматах). При paranoiac аномально усилены реакции избегания. Анализ физиологических процессов показал, что эти мутации нарушают нормальную функцию мембраны и почти не влияют на реснички. Существуют данные об аномальных электрофизиологических реакциях, соответствующих нарушениям разных типов.
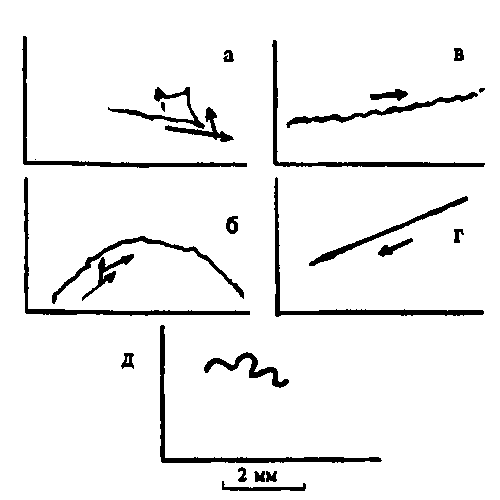
Рис.8.5. Фотография движения инфузории Раrаmесium: пройденный путь занял примерно 9 сек.
а — инфузория "дикого типа": видны спонтанные "реакции избегания" в виде изломов траектории; б — мутация fast-1, в — мутация pawn — пешка; г — paranoiac, д — sluggish — вялые животные
Сложность строения этой группы организмов, в особенности инфузорий, у которых функции целого организма осуществляются в пределах одной клетки, не позволяет считать их удобным объектом для исследования мутаций, влияющих на поведение. Однако они могут быть использованы для оценки эффектов фармакологических агентов.
Свободно живущая почвенная нематода Caenorhabditis elegans была выбрана в качестве объекта исследования С. Бреннером в начале 70-х годов (рис. 8.6, а). Длина этого червя составляет около 1 мм, его репродуктивный цикл длится 3,5 суток. C.elegans — гермафродит, размножается самооплодотворением, откладывает до 300 яиц. Иногда в потомстве появляются особи только с мужскими половыми органами, которые спариваются с гермафродитами. Этот тип размножения дает дополнительные возможности для генетических исследований. У этого вида в гаплоидном наборе 5 аутосом и X-хромосома. Построена почти полная карта генома C.elegans. По степени сложности строения тела и физиологических реакций эта нематода занимает промежуточное положение между Е.coli и дрозофилой. Геном C.elegans состоит из 8*10^6(?) пар оснований, что в 20 раз больше, чем у E.coli, и примерно в 2 раза меньше, чем у дрозофилы. Около 83% генома нематоды составляют уникальные последовательности.
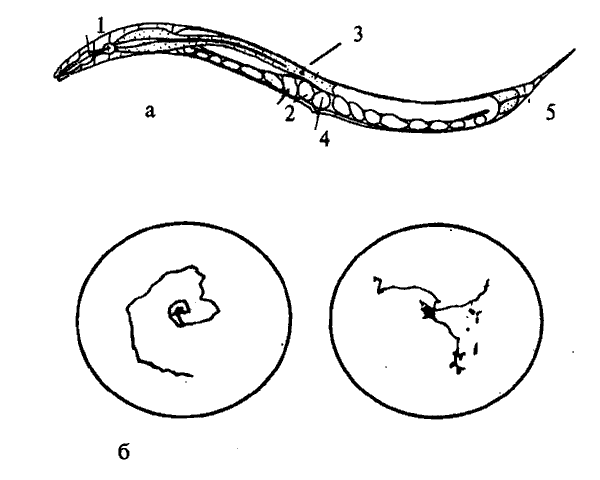
Рис.8.6. Относительно новый генетический объект: микроскопическая почвенная нематода Caenorhabditis elegans. а — схема анатомии червя: 1 — глотка; 2 — кишка; 3 — яйца; 4 — влагалище; 5 — анальное отверстие.
б — следы на питательной среде агар-агаре, сделанные нематодой дикого типа с нормальными движениями (слева) и двумя нематодами с мутацией bent—head, влияющей и на скорость движения, и на координацию мышц туловища
Для нематод характерна эвтелия, т.е. наличие в организме фиксированного числа клеток. Тело C.elegans состоит из 1000 клеток, причем 302 (по другим данным 350) из них — это нейроны. Генетическое, анатомическое и физиологическое исследование этого объекта, предпринятое С.Бреннером, имело несколько конкретных целей.
Во-первых, предполагалось создать пространственную картину связей всех клеток нервной системы с помощью электронномикроскопического анализа многих тысяч серийных срезов.
Во-вторых, определялась химическая природа синапсов и выявлялись мутационные изменения, влияющие на поведение животного, последние сопоставлялись с изменениями в "рисунке" соединения клеток и/или химизме синапсов.
Описано несколько сот мутаций С.elegans, и примерно 2/3 из них затрагивают поведение. В спектр поведенческих изменений входят нарушения координации движений червя (рис. 8.6,6), аномалии строения тела, которые влекут за собой аномальные движения, изменения хемотаксиса, нарушение откладки яиц.
Примерно у половины мутантов с нарушенным поведением были найдены отклонения в строении нервной системы. Их можно разделить на 4 категории.
1. Изменения в клетках-предшественницах нейронов. Поскольку ход нормального развития нейронов из нейробластов изучен у этого вида достаточно хорошо, мутации в этих элементах легко обнаружить. Они могут проявляться в виде особенностей поведения на разных стадиях развития организма.
2. Изменения в специфичности синаптических связей. Поскольку соединения нейронов взрослой особи дикого типа уже известны, возникающие вследствие мутаций структурные изменения синаптических соединений можно с достаточной надежностью сопоставить с аномалиями поведения у таких животных.
3. Изменения в расположении отростков нейронов. Созданная полная топографическая карта нервной системы С.elegans позволяет выделить таких мутантов, у которых отростки нейронов идут к аномальным мишеням. Такие мутации изменяют направление прорастания и характер контактов у нейронов сразу нескольких классов. Очевидно, что изменения поведения при этом достаточно разнообразны.
4. Изменения в функции нейромедиаторов. Эту категорию эффектов оказалось возможным проанализировать с развитием методов иммуноцитохимии и биохимии. Данные о медиаторной специфичности конкретных нейронов, как правило, оказываются неточными.
Электрофизиологические исследования этого объекта практически невозможны из-за его малого размера, поэтому сведения о нейрохимической специфичности нейронов того или иного класса сопоставляют с электрофизиологическими реакциями сходных групп клеток другой нематоды — Ascaris. У C.elegans обнаружены классические нейромедиаторы — ацетилхолин, ГАМК и биогенные амины.
Реакция на химические сигналы. Мутации, индуцированные химическими агентами, обычно бывают рецессивными и выявляются во втором поколении. Изучение поведения C.elegans и его изменений у мутантов показывает, что если у бактерий опознание химического вещества базируется на времени удержания следа, оставленного веществом на мембране и внутри клетки, то у C.elegans все обстоит сложнее. Так, например, мутант slow перемешается в 8 раз медленнее, чем особи дикого типа, но отыскание химических веществ, необходимых для питания, происходит у него без отклонений. Таким образом, для этой реакции время сохранения следа значения не имеет. Сопоставление градиента концентрации химического вещества вдоль тела червя также не может служить основой такой способности, так как мутанты, у которых изменены хеморецепторы хвостовой части тела, тем не менее, могут нормально отыскивать вещества. По всей видимости, основой реакции на химические вещества служит клинотаксис, т.е. движения, возникающие в результате оценки различий концентрации вещества с двух сторон тела. Поведенческим проявлением клинотаксиса, как считается, могут быть так называемые "рыскающие" движения головного конца.
Локомоцня. Координация движений. В виду очень мелких размеров C.elegans основным методом выявления нарушений движения у него стала регистрация следов, которые червь оставляет на поверхности агара, служащего ему питательной средой. Примером мутаций, нарушающих движения, может служить roller, когда червь перемещается, переворачиваясь через головной и хвостовой концы, и bent head: при этой мутации изменена анатомия головного конца, и вместо обычных "рыскающих" движений перемещение идет по спирали. Движения червя обеспечиваются элементами брюшной нервной цепочки. Ее нейроны и их соединения хорошо описаны, известны также и эмбриональные клетки-предшественницы этих нейронов. Выделяют пять классов нейронов, которые подразделяются на два главных типа: возбудительные нейроны, инициирующие движение животного вперед или назад, и тормозные нейроны, координирующие сокращения мышц двух сторон тела.
Мутации группы Unc (uncoordinated) были одними из первых, описанными у C.elegans. Аномалии движений при неврологических мутациях очень перспективны для исследования организации движений в норме. Фенотипически группа мутаций Unc подразделяется на 3 категории:
1) животные либо сохраняют неподвижность в покое, либо при стимуляции, которая в норме побуждает их двигаться вперед или назад, сокращаются все мышцы одновременно; такие системные мутации затрагивают функцию тормозных нейронов, в частности влияют на распределение их отростков (unc-30); в других случаях нарушается синтез ГАМК;
2) животные не способны двигаться в одном из направлений (вперед или назад), а при соответствующей стимуляции закручиваются либо дорсальной, либо вентральной стороной кверху; установлено, что в основе такого дефекта лежат аномальные связи интернейронов; в некоторых случаях отсутствует иннервация ряда мышц;
3) животные неспособны двигаться по прямой, не извиваясь; существуют свидетельства того, что у таких мутантов аномалия затрагивает холинергические синапсы.
Реакция откладки яиц. Выход яиц из организма обеспечивают мышцы, иннервация которых идет от двух категорий нервных клеток, по-видимому, серотонинергических. Показано также, что октопамин эту реакцию тормозит, а его антагонист стимулирует. Описано около 40 мутаций, изменяющих процесс. При одной из них, например, нарушено собственно выведение яиц, и молодые особи вылупляются, оставаясь внутри родительского организма.
Мутации, влияющие на продолжительность жизни. Продолжительность жизни C.elegans — 18-21 день. Этот объект очень удобен для исследования молекулярно-генетических основ старения. Еще в начале работы с C.elegans была описана мутация age-1, при которой продолжительность жизни увеличивается до 34 дней.
Еще одна группа мутаций, влияющих на продолжительность жизни, связана с особенностями жизненного цикла этой нематоды. По окончании эмбриогенеза из яйца выходит личинка первой стадии (L1), которая затем проходит фазы L2, L3 и L4, после чего особь становится половозрелой. Однако в жизненный цикл этой нематоды может включаться еще одна, специфическая стадия — стадия покоящейся личинки, называемой дауером (dauer) — стадия d. В таком состоянии животное успешно переживает неблагоприятные средовые условия, например перенаселение или недостаток пищи. Стадия d может наступить в период после стадии L2 вместо L3. При восстановлении пригодных для жизни внешних условий из стадии d червь переходит в стадию L4.
В настоящее время предполагается, что в стадии d у животного происходит своеобразное выключение процессов старения, поскольку подобные покоящиеся личинки могут оставаться жизнеспособными на срок до 1 года. Выявлено семейство генов daf, участвующих в формировании стадии покоящейся личинки. Манипулируя условиями выращивания личинок с мутацией гена daf-2, можно получить особей, которые почти столь же жизнеспособны, как и особи дикого типа, но длительность их жизни значительно увеличена — до 43 дней. По существующим данным, у долгоживущих мутантов, отмечаются биохимические особенности, возможно связанные с изменением интенсивности процессов старения.
Пластичность поведения. Из всех описанных поведенческих реакций этой нематоды наиболее полно изучена реакция избегания в ответ на прикосновение. При стимуляции хвостового конца тела волоском червь реагирует продвижением вперед, при прикосновении к головному концу пятится назад. Неврологическим субстратом этих реакций является группа из 6 тактильных рецепторов, 5 пар интернейронов и 69 мотонейронов. Сочетание данных мутационного анализа и исследования фенотипа ряда мутантных форм (всего описано несколько сот таких мутаций) с микрохирургическими вмешательствами в ЦНС червя с помощью лазерного луча позволило установить подробную схему соединений нервных элементов этой сети. Важно отметить, что эффект мутаций проявляется всегда в виде целого набора морфофункциональных изменений, и даже внимательный поиск не обнаруживает мутаций, эффект которых проявлялся бы в аномалии какого-то одного элемента этой сети.
Несмотря на относительную простоту, поведение C.elegans может видоизменяться в результате накопления индивидуального опыта. Экспериментально показано, что у этих животных могут происходить процессы: неассоциативного обучения — привыкание к регулярно действующему стимулу (механическое сотрясение), растормаживание привыкания и сенситизация. Каждая из личиночных стадий имеет специфические особенности в формировании привыкания к тактильным стимулам. Обнаружение в будущем мутаций, влияющих на процессы неассоциативного обучения, в сочетании с беспримерным уровнем изученности морфофункциональных основ поведения этих животных даст новое понимание механизма пластичности реакций нервной системы (подробнее см.: Rankin, Best, 1992).
Относительно новый генетический объект, каким является C.elegans, очень удобен для изучения молекулярных основ патологии человека. В рамках большой исследовательской программы по генетическим механизмам болезни Альцгеймера были проведены эксперименты с получением трансгенных нематод. Как известно, одним из типичных проявлений патологии при болезни Альцгеймера являются отложения так называемого бета-амилоидного белка в виде бляшек.
У трансгенных особей C.elegans ген unc-54, связанный с функцией мышц, был соединен с последовательностью оснований геном, кодирующим бета-амилоидный пептид. Этот чужеродный для нематоды ген экспрессировался, и его продукт был обнаружен. Анализ иммунореактивности в тканях таких генетически видоизмененных организмов позволил найти отложения, сходные с классическими бляшками бета-амилоидного белка, характерными для болезни Альцгеймера (см.: 8.6.5.5).
Таким образом, микроскопическая нематода как объект генетики поведения не только дает возможность исследовать нейроморфологические основы примитивных реакций, но и становится удобным объектом для исследований по перенесению генетического материала (генной инженерии).
Аплизия, или морской заяц (Apfysia californica), — объект, на котором проведены классические эксперименты по физиологии поведения моллюсков. В них продемонстрирована роль индивидуальных нервных клеток (так называемых индентифицированных нейронов) в формировании поведенческих реакций организма. Это новое направление — физиология поведения животных с "простыми нервными системами" (см.: Кенделл, 1980).
Исследование нейронных механизмов физиологических процессов у этого брюхоногого моллюска, например особенностей гормональной регуляции реакции откладки яиц, для которой известны все ее этапы, позволило перейти на молекулярно-генетический уровень и установить генную регуляцию группы конкретных ФКД из репертуара поведения этого животного.
Откладка яиц у аплизии обеспечивается цепью действий, включающую торможение локомоции, усиление дыхательных движений, специфические движения головы, с помощью которых жгут с яйцами вытягивается, сворачивается и приклеивается к субстрату. Эти события происходят вследствие активации сетей нейронов, которая является результатом координированного выделения нескольких пептидов. Пептиды, выделяющиеся железами репродуктивного тракта после копуляции, возбуждают нейросекреторные "пазушные" клетки. Они, в свою очередь, выделяют набор пептидов, которые воздействуют на разные нейроны моллюска. Возбуждение (или модуляция работы) пазушных клеток запускает сложную последовательность действий аплизии. Была выделена последовательность ДНК, кодирующая белок-предшественник, содержащий аминокислотные последовательности упомянутых пептидов. Считается установленным, что за формирование набора химических агентов, запускающих ФКД откладки яиц аплизии, отвечает один ген.
Сердечно-сосудистые реакции аплизии исследованы достаточно подробно в аспекте анализа функции пептидов (и их белков-предшественников), запускающих соответствующие нейроны. Пептид с последовательностью аминокислот Phe-Met-Arg-Phe, так называемый FMPF-амид,, описанный впервые у моллюсков, известен тем, что иммунореактивность к нему обнаруживается у всех представителей животного царства.
Генетика аплизии в классическом понимании не изучена: данные о генетической изменчивости особей, о выявленных мутациях и даже о строении ее генома пока отсутствуют. В то же время исследования, проведенные на этом моллюске, оказали мощное влияние на формирование современных нейрогенетических представлений не только в области генетики поведения дрозофилы, как наиболее генетически изученного экспериментального животного, но и для нейрогенетики в целом (подробнее см.: Nambu, Kirk, 1992).
Медоносная пчела Apis mellifera — социальное насекомое, живущее большими семьями. Единственную плодовитую самку колонии — пчелиную матку — можно искусственно оплодотворить, используя сперму трутня нужного генотипа. Как известно, в размножении этих перепончатокрылых бесплодные рабочие особи колонии имеют 75% общих генов. В связи с таким способом размножения оценку генетической изменчивости пчел проводят, беря за единицу для анализа не отдельную особь, а пчелиную семью.
Классическая работа по генетике поведения пчел была проведена Н. Ротенбулером несколько десятилетий назад (Rothenbuhler, 1967).
Пчеловодам было известно, что пчелы некоторых пород устойчивы к инфекции: не болеют американской гнильцой. Породы, устойчивые к этому заболеванию, имеют к своем поведенческом репертуаре специфические "гигиенические реакции". Пчелы каким-то образом обнаруживают ячейки, в которых находятся инфицированные личинки, и выбрасывают их из улья, препятствуя тем самым распространению болезни.
Генетический анализ — оценка поведения гибридов, полученных от скрещивания устойчивых к болезни и подверженных заболеванию пчел, показал, что это "гигиеническое поведение" детерминировано двумя независимыми генами, причем аллели, определяющие устойчивость к заболеванию, рецессивны (рис. 8.7).
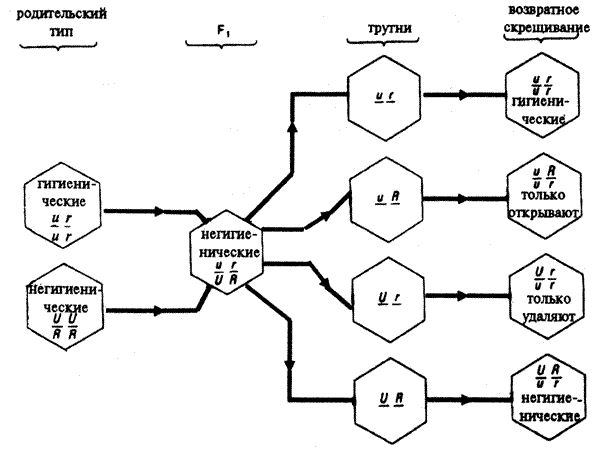
Рис.8.7. Схема наследования признака "гигиеническое поведение" у медоносной пчелы (см. текст)
Полученные в этом элегантном исследовании данные и сегодня остаются одним из немногочисленных примеров олигогенного наследования признаков поведения. Более подробно эти опыты изложены в учебнике Д.Мак-Фарленда (1988).
Движения пчел, которые они совершают при вскрытии ячеек и выбрасывании зараженных личинок, относятся к категории ФКД и обладают всеми характерными для них признаками. Данные движения выполняются насекомыми в полной форме без предварительного опыта и практически неизвестно, чтобы они видоизменялись при "тренировке". Более того, эти движения неспецифичны и характерны для вида в целом, а не только для каких-либо пород пчел. Все пчелы выполняют эти движения, однако у пород, подверженных заболеванию, частота этих ФКД чрезвычайно низка. Поэтому их поведение не может играть своей адаптивной роли: зараженные личинки остаются, и инфекция распространяется.
Генетические исследования фуражировочного поведения медоносной пчелы были проведены на селектированных линиях. Селекцию проводили по признаку "большое или малое количество собираемой пыльцы". После получения пятого поколения селекции провели скрещивание двух линий и проанализировали поведение гибридов F1 и F2, а также беккроссов.
Помимо классического генетического анализа был также использован метод картирования "локусов количественных признаков" (см.: 8.6.6.4), позволяющий с помощью специальных молекулярных маркеров выявлять гены, участвующие в определении данного признака, т.е. находить их положение на той или иной хромосоме.
Оказалось, что количество пищи, собираемое пчелой, детерминировано двумя локусами — pln-1 и pln-2 — в группах сцепления II и X.
Генетические данные, полученные как характеристика особенностей фуражировочного поведения колоний, были подтверждены при исследовании поведения отдельных особей. Особи-фуражиры из линии с высокими показателями сбора пищи всегда приносили больше нектара или пыльцы, чем особи из линии с низкими значениями признака.
Генетика падальной мухи (Phormia regina) изучена очень слабо, в особенности по сравнению с дрозофилой. В то же время крупные размеры этого насекомого позволили Дж.Хиршу (J.Hirsch) и его коллегам еще в 70-е годы провести на этом объекте исследования по генетике обучения пищевой реакции, которые и по сей день остаются уникальными (Mc Guire, Hirsoh, 1977).
Спомощью специальной пипетки насекомое фиксируется так, чтобы можно было наблюдать, как оно вытягивает хоботок (рис. 8.8).
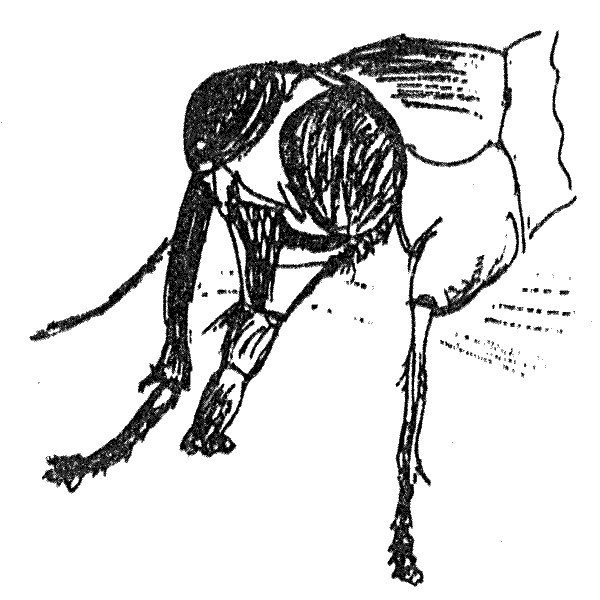
Рис. 8.8. Голова падальной мухи с хоботком, который вытягивается при попадании на вкусовые рецепторы сладкого раствора
Безусловной реакцией, на основе которой вырабатывается условный рефлекс, является вытягивание хоботка при нанесении на оральную область 0,5М раствора сахарозы. Нижние членики передней лапки, где расположены хемочувствительные нервные окончания, помещают либо в дистиллированную воду, либо в раствор какого-либо вещества (но не сахара). При такой схеме эксперимента у насекомого вырабатывается условная реакция вытягивания хоботка в ответ на хемосигналы, ранее не вызывавшие такой реакции. Однако, как это всегда бывает при развитии процесса обучения у примитивных животных, одновременно выявляется сильное влияние процесса сенситизации, т.е. изменения чувствительности эффекторной системы не за счет формирования ассоциативной связи, а как результат совместного действия ряда стимулов и возбуждения насекомого в результате фиксации. При исследовании пищевых реакций Phormia regina процесс сенситизации исследователи назвали центральным возбужденным состоянием (central exitatory state — CES).
Чтобы отделить феномен формирования истинной ассоциативной связи от явлений CES, была разработана специальная схема эксперимента.
Стимулы. УС1 + УС2 + БР
Время действия. 4с 4с 1 с + 10 мин перерыва
Действие: изолированное совместное
1 сек
При такой последовательности стимулов двигательная безусловная реакция (вытягивание хоботка) в ответ на УС1 является отражением CES, т.е. следствием увеличения чувствительности к безусловному стимулу. Реакция же на УС2 отражает формирование условной связи; 10-минутный перерыв после сочетания стимулов способствовал угасанию CES. К 15-му применению комплекса стимулов доля мух с условной реакцией росла, что свидетельствовало о формировании ассоциативной связи.
С помощью искусственного отбора на быстрое и медленное образование ассоциативной связи получили две линии мух, различия между которыми по исследуемому признаку постепенно увеличивались в процессе селекции. Искусственный отбор на быстроту формирования CES дал иную картину динамики различий: уже после первого поколения отбора селектируемые линии стали резко различаться по быстроте формирования CES, однако дальше по ходу селекции эти различия оставались неизменными (рис. 8.9).
Рис 8.9. Ответ на отбор а — по признаку "быстрое" или "медленное" формирование классического условного рефлекса (вытягивание хоботка при сочетании условного и безусловного вкусовых стимулов), б — на быстрое или медленное формирование ''центрального возбужденного состояния'' (CES)
Столь разные ответы на отбор у двух признаков свидетельствуют о разном характере их генетической детерминации. Быстрый ответ с последующим параллельным ходом кривых (как в случае CES) свидетельствует о моногенном наследовании признака, тогда как постепенное расхождение кривых, соответствующих уровням признака в разных поколениях, — следствие его полигенной детерминации.
Поскольку падальная муха — достаточно крупный объект, который позволяет вести нейрофизиологические исследования с меньшими трудностями, чем, например, на дрозофиле, результаты селекционных экспериментов представляются особенно интересными для изучения нейробиологии классического условного рефлекса.
Исследование генетических закономерностей в проявлении физиологических признаков у "простых" объектов, информация о дифференциальной экспрессии нейрогенов в корреляции с разными функциями существенно помогают в формировании новых подходов к анализу генетики поведения у более сложных, но традиционных объектов — дрозофилы и лабораторных грызунов.
Характерной чертой поведения насекомых (в том числе мух рода Drosophila) является преобладание в нем видоспецифических фиксированных комплексов действии. Однако наряду с ними в поведенческом репертуаре плодовой мушки имеются как более простые (таксисы), так и более сложные (реакции, приобретенные при обучении) феномены. Наиболее подробно изучено поведение Drosophila melanogaster.
Биологические ритмы. Характерный для дрозофилы ритм суточной активности обнаруживается не столько в уровне локомоции, сколько в частоте вылупления взрослых особей из куколок. Такие изменения в физиологических функциях очевидным образом модулируются чередованием света и темноты, однако, как это показано и для других животных, существуют также и внутренние механизмы поддержания околосуточных колебаний.
Фото-, гео- и другие таксисы. Это наиболее четкие, относительно примитивные реакции, выражающиеся в виде упорного перемещения насекомых в градиенте интенсивности света, концентрации химического вещества, геомагнитного поля и др. На стремлении мух перемещаться к свету (фототаксис) базируются методики исследования способности дрозофилы обучаться избегать неприятного запаха при его сочетании с ударом электрического тока.
Пищевое поведение. В ответ на вкусовое раздражение у мухи вытягивается хоботок. Эта безусловная реакция легла в основу выработки пищевого условного рефлекса у падальной мухи. Проведение таких исследований у плодовой мушки затруднено в связи с ее малыми размерами, однако известно, например, что эта реакция сильно различается у самцов и самок. Более плодотворны проводимые на дрозофиле исследования генетического контроля пищевого предпочтения. Эту особенность поведения изучают как на личинках (поиск пищи), так и на взрослых особях в связи с выбором самкой пищевого объекта для откладки яиц.
Реакцна обонятельные раздражители. Особый аспект пищевого поведения мух представляет собой их положительная реакция на обонятельные сигналы — пищевые аттрактанты. Подобные реакции осуществляются по механизмам таксисов, однако конкретных данных об их физиологических механизмах немного. Эти таксисы достаточно часто используются в экспериментах по генетике поведения, для которых такая информация необходима. В частности, положительная реакция на пищевой аттрактант не обязательно бывает связана с высоким уровнем локомоторной активности.
Оборонительное поведение. Обонятельные сигналы могут вызывать и реакции избегания. К их числу относится бензальдегид, широко используемый в экспериментах с обучением дрозофилы. В качестве отрицательного воздействия, на основе которого можно сформировать условную реакцию избегания, используется раздражение электрическим током. Физиологические корреляты такого воздействия изучены мало.
Половое поведение. У двукрылых, и в частности у дрозофилы, этот вид поведения организован достаточно сложно. Собственно спариванию предшествует более или менее интенсивно выраженное поведение ухаживания, функция которого — активация обоих партнеров (см.: рис. 8.1). Процесс ухаживания включает в себя несколько стадий. Первая из них — это стадия ориентации, когда самец занимает перед самкой положение, перпендикулярное продольной оси ее тела. Во время второй стадии он осуществляет вибрацию крылом, повернутым в горизонтальной плоскости на 90o. Эта фаза очень вариабельна по длительности (она укорочена при мутации yellow) и по "рисунку" отдельных пачек сигналов, а также интервалов между ними. Затем следуют стадии облизывания гениталий и собственно садки.
После копуляции у самки развивается временная половая невосприимчивость, а в ее организме образуется химическое вещество, вызывающее у самцов реакцию избегания. Это специфическое вещество цис-вакцинилацетат называется антиафродитным феромоном. Генетические исследования полового поведения некоторых мутантов (см.: 8.4.3.5) показали, что самец, который сделал неудачную попытку ухаживания за невосприимчивой самкой, в дальнейшем, в течение примерно 2 ч воздерживается от ухаживания даже за рецептивными самками. Иными словами, мухи обнаруживают способность к торможению поведенческой реакции вследствие неприятного раздражения, т.е. они обучаются после единственного сочетания условного (вид самки) и безусловного (антиафродитный феромон) сигналов.
Обучение. Способность дрозофилы к обучению реакции избегания электрического тока исследуется в специальных установках с помощью методики "обратного потока" (см.: 8.4.3.6). Этот метод позволяет быстро тестировать большое число особей, что особенно важно при поиске мутантов с измененным поведением.
Очевидное адаптивное значение в жизни дрозофилы в естественных условиях имеет недавно описанная способность этих насекомых обучаться подавлению локомоторной активности. В экспериментах, проведенных в Институте физиологии им.Павлова РАН Н.Г. Камышевым с соавт. (1994), четко продемонстрировано, что исходный уровень локомоторной активности мухи достоверно снижается, когда ее подсаживают к группе из 10 других мух, причем, это снижение нельзя объяснить ни реакцией на какие-либо вещества, ни безусловнорефлекторным торможением активности, ни привыканием.
Снижение двигательной активности мух выражается, в частности, в укорочении длительности отдельных "пробежек" и снижении числа контактов особей между собой. Оно имеет условнорефлекторную природу и является результатом формирования инструментального навыка подавления активности в результате отрицательного опыта: при контактах мухи обмениваются "ударами" конечностей, что играет роль наказания. В естественных условиях, при скоплении дрозофил на источнике пищи — гниющих фруктах — способность к подобному обучению, видимо, обеспечивает оптимальное использование пищевых ресурсов, а это, несомненно, способствует выживанию.
Первоначальное накопление информации по генетике поведения дрозофилы происходило при исследовании плейотропных эффектов отдельных мутаций, влияющих на цвет глаз и тела. Выше (см.: 8.2.2) уже упоминалось о плейотропном эффекте мутации yellow, когда у самцов достоверно снижается способность к ухаживанию: у них сильно укорочена во времени фаза "вибрации". Эти особи проигрывают в ситуации конкуренции с самцами других генотипов, однако в целом линии с мутацией yellow размножаются в нормальном темпе за счет увеличенной половой активности самок.
Ген ebony, который изменяет цвет хитиновых покровов мухи, также оказывает плейотропный эффект на ее поведение. Эта мутация сопровождается изменением содержания дофамина в мозге и изменением двигательной активности мух, что в свою очередь сказывается на уровне половой активности обоих партнеров.
При мутации black в организме дрозофилы не синтезируется бета-аланин, хотя его усвоение возможно. Введение бета-аланина только что вылупившимся мухам предотвращает плейотропные эффекты этой мутации. Такие особи фенотипически нормальны.
В 70—80-е годы в Институте физиологии им.Павлова АН СССР было проведено обширное исследование эффектов мутаций медоносной пчелы и дрозофилы, нарушающих метаболические превращения триптофана в той его, части, которая связана с синтезом так называемых кинуренинов — веществ, непосредственно участвующих в образовании пигментов глаз насекомых (Пономаренко и др., 1975).
Ниже схематически представлены основные этапы этого обмена с указанием места возникновения дефекта (т.е. аномалии обмена) при разных мутациях у представителя отряда двукрылых — Drosophila melanogaster и у представителя перепончатокрылых — Apis mellifera. Стрелкой показаны участки цепи метаболических реакций, которые страдают при соответствующих мутациях.
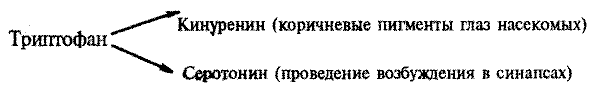
Физиологический анализ эффектов этих мутаций у обоих видов показал, что эффекты сходны между собой и практически параллельны у дрозофилы и у пчелы, и что в целом они связаны с модуляцией уровня возбудимости ЦНС, в частности уровня двигательной активности.
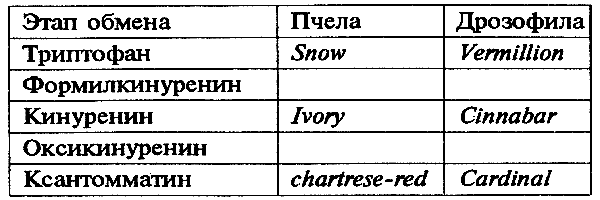
Мутация chartrese-red пчелы, например, сопровождается изменениями возбудимости нервно-мышечного аппарата и ритма танца и накоплением в организме промежуточного метаболита З'-гидроксикинуренина. У дрозофилы эти мутации вызывают модуляцию уровня двигательной активности, увеличение длительности наркоза при действии эфира, снижение возбудимости нервно-мышечного аппарата.
Физиолого-биохимическое исследование показало, что основной причиной сходства эффектов таких мутаций является накопление в организме насекомых обоих видов избыточных количеств серотонина. Эта интересная генетическая модель показывает, насколько сложными и неожиданными могут быть поведенческие эффекты мутаций, которые внешне выражаются всего лишь в изменении цвета глаз.
Для анализа общебиологических закономерностей контроля жизнедеятельности не только насекомых, но и других групп животных представляют интерес генетические изменения суточного ритма. Исследуются, например, мутации дрозофилы, изменяющие длительность циркадного (суточного) ритма уровня двигательной активности мух и ритма массового вылупления взрослой формы из куколки. При мутации в локусе arythmic суточные колебания активности отсутствуют; мутации в локусе period, либо укорачивают цикл активности (short period) до примерно 19 ч, либо удлиняют его (long period) до примерно 28 ч. При мутации per 0 суточный ритм практически отсутствует.
У разных видов дрозофилы описано около 10 мутаций, влияющих на суточный ритм. Они обнаруживаются как в их воздействии на суточные колебания локомоторной активности, так и в модуляции периода массового вылета взрослых особей из куколок. Ритмичность физиологических процессов, влияющих на локомоцию и приуроченность вылупления к определенной фазе циркадного ритма, обусловлена согласованием световосприятия с работой эндогенного водителя ритма (пейсмейкера).
Это было показано при исследовании аутосомной рецессивной мутации linne, когда ритмичность вылупления при определенных условиях поддерживается, однако смена света и темноты не может запускать ритм.
Геотаксис. Первые систематические и подробные эксперименты по генетическому контролю геотаксиса у Dr. melanogaster провел в 60-е годы Дж.Хирш (см.: Эрман и Парсонс, 1984). Геотаксис, т.е. перемещение мух, ориентированное относительно силы тяжести, оценивали с помощью специально сконструированных лабиринтов (рис. 8.10).
Использовали вертикально расположенный лабиринт, представляющий собой систему соединенных друг с другом модулей, в каждом из которых муха может выбрать между поворотом в одну сторону, который ведет ее вверх, и поворотом в другую, при котором она спускается вниз. Обычно использовали 10- или 15-модульный лабиринт. Группу мух помещали в единственную пробирку, расположенную на стартовой стороне лабиринта, и давали им возможность перемещаться в направлении другой стороны лабиринта. Привлекательными стимулами были либо запах пищи, либо свет (рис. 8.10).
Рис. 8.10. Лабиринт для изучения геотаксиса у плодовой мушки.
Данная конструкция располагается вертикально, и муха, запущенная на "старте" до того, как она доберется до "финиша", должна преодолеть некоторое число разветвлений лабиринта, в каждом из которых она может повернуть вверх (против силы тяжести) или вниз (в направлении силы тяжести). На финише мух собирают из ряда пробирок, причем в самой верхней оказываются насекомые с наиболее выраженным отрицательным геотаксисом, в нижней — с максимально развитым отрицательным фототаксисом Отдельно показано устройство звена-разветвления: сужение перед поворотом не позволяет мухе вернуться назад
Поскольку на финишной стороне лабиринта находится не одна, а 10 или 15 пробирок, расположенных на разной высоте по отношению к стартовой, то происходит сортировка мух по уровню проявления их геотаксиса. Такая методика позволяет провести отбор на положительный и отрицательный геотаксис у дрозофилы (рис. 8.11), причем селекция происходит очень быстро. В дальнейшем было проведено аналитическое исследование роли "вклада" генов каждой из хромосом дрозофилы (за исключением 4-й) в обеспечение отрицательного и положительного геотаксиса. Это оказалось возможным благодаря использованию специальных линий, которые несли доминантные маркеры, а также инверсии, препятствовавшие кроссинговеру определенных хромосом. Была получена количественная характеристика линий дрозофилы, селектированных на разные уровни геотаксиса, и определена степень участия разных хромосом в детерминации этого признака (подробнее см.: Эрман, Пар-сонс, 1984).
рис.8.11. Уровень геотаксиса у трех линий дрозофилы — контрольной (неселектированной) линии и линий, селектированных соответственно на отрицательный и положительный геотаксис. По оси ординат — доля животных (в процентax), обнаруживших тот или иной индекс геотаксиса
Поведение мух, селектированных на отрицательный геотаксис в течение 181 поколения, имело целый ряд особенностей, привнесенных процедурой тестирования в лабиринте. Оказалось, что параллельно с отбором на крайние значения геотаксиса эти мухи подвергались неосознанному отбору: на устойчивость к высыханию и способность переносить длительные периоды голодания, поскольку перемещение некоторых мух занимало до нескольких часов и многие из них погибали; на сниженный уровень двигательной активности, поскольку более активные насекомые имели тенденцию к гибели; на усиление стереотипии в поведенческих реакциях, поскольку мухи, обладающие крайними значениями геотаксиса, имели тенденцию делать повороты в одну и ту же сторону, в том числе и в лабиринте, который был расположен горизонтально; на сниженный уровень "клаустрофобии", поскольку мухам приходилось преодолевать суженные участки пробирок при переходе из одного блока лабиринта в другой и некоторые насекомые не справлялись с этим.
Результаты экспериментов показывали сложность поведения дророфилы и возможность формирования иных, чем предполагалось изначально, ответов на отбор. Подобное явление относится к так называемым коррелированным ответам, достаточно подробно изученным также в экспериментах по генетике поведения грызунов (см.: 8.6.3.1).
Фототаксис. Следующий этап в накоплении информации о генетических основах поведения дрозофилы связан с именем американского ученого С.Бензера (1975), в работах которого был использован целый комплекс экспериментальных подходов.
В частности, был активизирован поиск неврологических мутантов, и с этой целью были разработаны специальные приемы массового тестирования мух.
Отбор потенциальных носителей аномалий проводили с помощью метода обратного потока (рис. 8.12). По выражению Бензера, этот способ позволял сортировать мух, как молекулы. Сущность его заключалась в следующем. Разделяли мух, которые двигались внутри трубки в направлении источника света, и тех, которые оставались на месте. Процедура повторялась несколько раз. В результате была получена серия групп насекомых, у которых выраженность исследуемого признака варьировала от минимального до максимального значения.
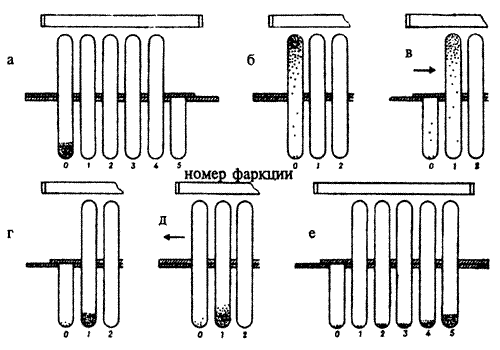
Рис. 8.12. Прибор для "сортировки" мух методом обратного потока. Аппарат состоит из двух групп пробирок, укрепленных в двух штативах, которые можно сдвигать относительно друг друга. Мух помещают в пробирку 0, после этого аппарат ставят вертикально и легким постукиванием сбрасывают их на дно пробирки. После этого всю систему кладут плашмя, так чтобы верхние концы пробирок были обращены к свету (а). Мухи, обладающие фототаксисом, движутся к свету, а другие остаются в темном конце (б). Через 15 с обращенный к свету (на рисунке — верхний) ряд пробирок сдвигают вправо (в) и снова стряхивают мух вниз (г). При этом мухи, реагировавшие на свет, попадают в пробирку 1. "Верхний" ряд пробирок сдвигают влево (д) и после перемещения системы в горизонтальное положение дают возможность мухам, проявившим фототаксис, снова бежать к свету. Процедуру повторяют 5 раз. В результате наиболее активные мухи оказываются в пробирке 5, несколько менее реактивные — в пробирке 4 и т.д. (е)
На следующем этапе из числа мух с положительным фототаксисом следовало отделить более активных особей от тех, которые энергичнее шли именно на свет. Мух с отрицательным фототаксисом также разделяли на малоактивных и на тех, которые избегали света. С этой целью мухам предлагалось двигаться в трубке от источника света. В результате можно было получить 5 групп насекомых, 4 из которых попарно различались либо по знаку фототаксиса, либо по уровню активности, у 5-й группы фототаксис отсутствовал (они не реагировали на свет) и были малоактивными (рис. 8,13). Полученные группы насекомых, достоверно различавшиеся по поведению, можно было исследовать нейрофизиологически. Запись электрических потенциалов глаза показала, что у части особей электроретинограмма отсутствовала. В ряде случаев у молодых мух электрическая реакция глаза на свет была нормальной, но затем происходила дегенерация фоторецепторов.
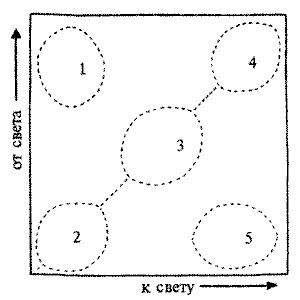
Рис. 8.13. Схематическое расположение пяти групп мух, отсортированных методом обратного потока (см. рис. 8.12) по разному уровню активности и разному уровню (и знаку) фототаксиса. 1 — мухи с отрицательным фототаксисом; 2 — медлительные мухи; 3 — мухи без фототаксиса и с невысокой активностью; 4 — "бегуны"; 5 — мухи с нормальным положительным фототаксисом
Особенности формирования аномального фенотипа у мутантов с нарушенным зрением были исследованы в экспериментах с гинандроморфными мозаиками (см.: 8.5.3.2). Первой в этом ряду была классическая работа А.Хотта и С.Бензера, кратко изложенная выше.
Проявление признака — аномальные зрительные реакции — исследовали у особей, часть клеток которых не имела Х-хромосомы. В таких клетках (они развились по "самцовому" типу) обнаруживался эффект упомянутых выше рецессивных генов, влиявших на фенотип.
Судорожные состояния. Еще одна группа аномалий с мутациями в Х-хромосоме была обнаружена случайно при наркотизации мух в ходе очередной серии опытов. Оказалось, что у этих особей при действии эфирного наркоза развиваются быстрые подрагивания конечностей или вспышки некоординированных движений, которые распространяются среди группы мух наподобие цепной реакции. Наиболее полно описаны мутации shaker (sh) и hyperkinetic (hk), в частности выявлены аллели гена hk, при которых мухи резко подпрыгивают и падают на спину при внезапном появлении в поле зрения движущегося предмета.
В знаменитых экспериментах американских ученых К.Икеды и В.Каплана в начале 70-х годов с помощью метода гинандроморфных мозаиков был прослежен путь проявления таких аномалий. Анализ физиологических процессов у особей, ткани которых представляли собой "мозаику" из участков нормального и мутантного генотипов, показал, что подергивание лапок при эфирном наркозе наблюдается не у всех мух. Мотонейроны, отвечающие за движения конечностей, располагаются в торакальном ганглии. Если ткань этого ганглия оказывается полностью или частично мутантной, то движения всех или части конечностей аномальны. Если же ткань торакального ганглия не несла мутантного аллеля, то аномальных подергиваний конечностей не было, даже если выше расположенные участки ЦНС (надглоточный ганглий) были мутантными. Таким образом, для проявления эффекта мутации принципиально важным оказывается экспрессия мутантного аллеля в нервных клетках, непосредственно проецирующихся в мышцы (рис. 8.14).
Рис. 8.14. Типы мозаичности фенотипа у гинандроморфных особей D.melanogaster по мутации hyperkinetic.
Ткани самца светлые, ткани самки темные, s — дрожащие лапки, в области их проекции в грудном ганглии обнаружены аномальные разряды, а — половина тела мухи нормальная, другая половина мутантная; б, в — мутантные ткани почти всего тела мутации сисирпопу.
У большинства видов животных собственно копуляции предшествует ритуал ухаживания, в процессе которого партнеры по очереди осуществляют видоспецифические движения (ФКД). Половое поведение дрозофилы намного примитивнее, чем у птиц и млекопитающих, коммуникационные процессы которых трудны для исследования именно из-за их сложности. У дрозофилы простота и четкость взаимодействии партнеров при спаривании видоизменяются в результате некоторых мутаций, что позволяет уточнить сложную мозаику их чередования.
Уже упоминалось о влиянии мутации yellow на фазу вибрации при ухаживании самца: она оказывается укороченной, и уровень стимуляции самки, необходимый для продолжения ритуала ухаживания, достигается только потому, что возбудимость самок этой линии выше, чем у самок дикого типа. Нарушение нормального ритма вибрации происходит и при мутации cacophony.
Частота биений крыла, длительность отдельных "пульсов" и интервалов между ними — это признаки, по которым различаются близкие виды, обеспечивая межвидовую изоляцию. Так, например, виды-двойники — Dr.simulans и Dr.jacuba — различаются "рисунком" видовой "песни"-вибрации. Поскольку движения крыла при вибрации — это пример примитивного инстинктивного движения, по своим свойствам относящегося к ФКД, генетические различия подобного рода подтверждают предположение о существовании резерва генетической изменчивости, обеспечивающего варьирование частоты выполнения ФКД (см.: 8.2.5).
Генетический контроль ухаживания очевидным (а может быть, и тривиальным) образом связан с процессом половой дифференцировки, поскольку мутации, затрагивающие последнюю, влияют и на поведение ухаживания. Выше упоминалось о том, что после спаривания в половых путях самки образуется специальное сигнальное вещество — антиафродитный феромон цис-вакцинилацетат. Процесс его формирования сложный. Оказалось, что образование сигнальных веществ этой группы связано с генотипом самца, спаривание с которым было успешно завершено. В сперме самца присутствует специфический фермент из группы эстераз, конкретная изоферментная форма которого детерминируется определенным аллелем данного локуса.
Именно его каталитическая активность в половых путях самки продуцирует (или не продуцирует) цис-вакцинилацетат.
Роль поведения самки в осуществлении самцом ритуала ухаживания была проанализирована еще в 70-е годы с использованием метода гинандроморфных мозаиков (см.: 8.5.3.2). Половое поведение мозаичных по "составу" своего тела мух, у которых доли "самцовых" и "самочьих" тканей различались, в разных условиях варьировало. Однако для процедуры простого преобладания "самочьих" черт строения было недостаточно. Критическими в этом отношении оказались определенные участки тела, а именно спинные хитиновые щитки — тегриты, щитки у основания ног — стерниты, а также область грудного интегумента (покрова). Когда эти участки имели генотип самки, то, даже если по другим морфологическим признакам муха была похожа на самца, она вызывала у самцов четкую реакцию ухаживания. Было показано, что ухаживание, в частности фаза вибрации, запускается обонятельными ключевыми раздражителями (феромонами), источником которых и являются указанные участки тела.
Исследование полового поведения гинандроморфных мозаиков показало, что эффекты разных мутаций определяются изменениями в разных участках ЦНС мухи. Отставление крыла и ориентация самца относительно самки, нарушающиеся при некоторых мутациях, связаны с работой надглоточного ганглия. Она повреждается при мутациях fruitless и celibate, и это ведет к нарушению копуляции. Изменение видоспецифической "песни"-вибрации при мутации cacophony связано с дефектом в торакальном ганглии мухи. О сложности механизма ритуала ухаживания говорит и тот факт, что он изменяется при мутациях в локусе per, когда нарушается и циркадная ритмика ряда процессов (см. выше). Возможно, что физиологические механизмы, обеспечивающие ритмические осцилляции, и механизмы, координирующие последовательности действий при ухаживании, либо управляются из одного отдела ЦНС, либо связаны с функцией одной медиаторной системы (рис. 8.15).
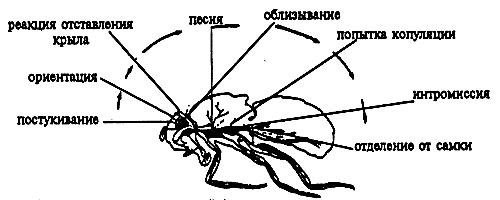
Рис. 8.15. Гены, которые в мутантной форме вызывают изменения показанных на рисунке поведенческих реакций; их экспрессия в ЦНС
Для массового тестирования поведения мух широко используется метод "обратного потока" (см. рис. 8.12). Несложное техническое устройство, которое позволяет "фракционировать" мух с разной выраженностью соответствующего признака, используется практически во всех исследованиях, связанных с обучением дрозофилы (рис. 8.16).
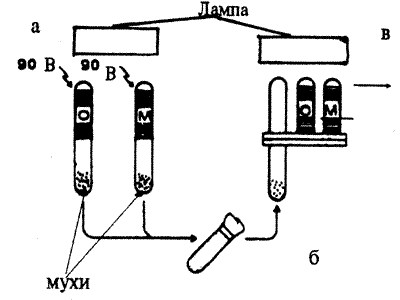
Рис. 8.16. Выработка реакции избегания у дрозофилы.
а — обучение: мухи, помещенные в пробирку, двигаются в направлении источника света и подвергаются действию удара тока в присутствии одного (О) или другого (М) пахучего вещества. Через некоторое время (б) мух тестируют (в): оценивают их реакцию избегания (по подавлению фототаксиса) на вещества О и М в отсутствие удара тока
Группу из 30—50 мух помещают в специальную трубку, внутри которой они могут перемещаться в направлении, например, источника света. В тестовую трубку подают пары вещества, обладающего сильным, но индифферентным для них запахом. Продувание паров пахучего вещества сопровождается подачей электрического тока на специальные металлические пластины-электроды, протянутые вдоль всей трубки. Включение тока вызывает у мух оборонительные реакции. По прошествии определенного интервала времени их можно тестировать повторно, предлагая на этот раз только запах, но не включая электрический ток. При этом регистрируют число мух, которые все-таки переместились в направлении света при предъявлении условного (запахового) сигнала, несмотря на то что ранее она сопровождалась неприятным стимулом. Это число мух сопоставляется с данными по той же группе насекомых, когда они просто перемещались в градиенте нового для них, но также нейтрального запаха.
Достоверность различий в реакциях на запах с "наказанием" и на истинно нейтральный запах (получавшие удар тока мухи двигались к свету медленнее) свидетельствовала об имевшем место научении насекомых.
При описании эффектов наиболее известных мутаций, вызывающих аномалии обучения и памяти, таких как dunce (dnc), turnip (tur), cabbage (cab), rutabaga (rut), zuccini (zuc), локализованных в Х-хромосоме, было продемонстрировано, что процесс восприятия запаха у таких мух не изменен, т.е. сенсорная функция не нарушена, следовательно, дефект связан с нарушением других функций ЦНС.
При мутации dnc резко сокращается время существования следа краткосрочной памяти. Через 10—15 секунд после сочетания стимулов эти мухи уже "не помнят", что получили наказание. У мутантов rut изменена не только краткосрочная память, но страдает и сам процесс обучения. Мутации tur и amnesiac (amn) также характеризуются сильно нарушенной памятью, хотя и в меньшей степени, чем dnc.
Полученные в экспериментах мутации, влияющие на процесс обучения реакции избегания удара тока и на хранение следов памяти, были в дальнейшем проанализированы с точки зрения изменений естественного поведения мух. Напомним, что в нормальных условиях самец дрозофилы делает попытку ухаживать за любыми самками, в том числе и за невосприимчивыми к ухаживаниям после оплодотворения. Последние на попытку ухаживания отвечают, как уже отмечалось, специфической реакцией выделения антиафродитного феромона, что вызывает у самца длительное (1—2 ч) подавление готовности к спариванию даже с рецептивными самками. Оказалось, что самцы с мутацией dnc готовы ухаживать за любыми самками (и даже за самцами) сразу же после контакта с оплодотворенной самкой. Мухи с мутацией amn, при которой дефект памяти незначителен, "помнят" об опыте отрицательного взаимодействия несколько дольше. Мутация amn нарушает также готовность самок к спариванию (рис. 8.17).
Рис.8.17. Сохранение памяти у нормальных мух (1), а также у неврологических мутантов дрозофилы — amnesiac (2) и turnip (3) (гетерозиготы); 4 — выполнение навыка мухами, го-мозиготными по dunce и turnip (Мак-Фарленд, 1988)
Исследование нейрохимических коррелятов обучения и памяти у дрозофилы послужило мощным толчком к большой серии работ, посвященных роли генетического аппарата клетки в формировании следа памяти. На первых этапах такого исследования было выявлено, что данная группа мутационных изменений поведения характеризуется нарушениями в работе системы вторичных посредников, а именно в ферментной системе синтеза и деградации циклического аденозинмонофосфата, а также ферментных систем, связанных с последующим фосфорилированием белков.
Было показано, что ген dnc — это структурный ген фосфодиэстеразы II дрозофилы. Найдена гомология между нуклеотидной последовательностью этого гена и последовательностью, кодирующей белок-предшественник гормона отладки яиц аплизии (см.: 8.3.4). Этот факт интересно сопоставить с описанной для мутантов по dnc стерильностью самок, когда образование яиц сохраняется, но самка не может их отложить.
Далее было продемонстрировано, что при мутации в локусе rut нарушено свойство аденилатциклазы активировать кальций калмодулина. При мутации tur ослабляется активность протеинкиназы С, а также гуанилатциклазы с одновременным снижением числа рецепторов моноаминов. При мутации amn повышается уровень цАМФ.
Участие системы вторичных посредников в формировании следа памяти обнаружено у аплизии, дрозофилы и млекопитающих (у мышей). Общая схема влияния системы вторичных посредников на формирование памяти достаточно сложна, но очень упрощенно ее можно представить следующим образом. Внутриклеточный уровень цАМФ формируется вследствие активации аденилатциклазы под действием нейромедиатора. Циклический монофосфат, связываясь с протеинкиназой А, вызывает высвобождение каталитических субъединиц этого фермента, которые уже в ядре фосфорилируют так называемый белок CREB (cAMF response element binding protein) в положении Ser-133. CREB относится к категории регуляторных белков — транскрипционных факторов. В свою очередь он активирует ряд генов. Система активации CREB является звеном в цепи событий, участвующих в сохранении следа памяти в нервной системе аплизии, долговременной памяти дрозофилы (Yin et al, 1994) и мыши (Bourchaladze et al., 1994; см. также: 8.6.3.3).
При анализе генетического контроля развития нервной системы (нейрогенеза) исследователи концентрируют внимание на общих закономерностях процесса, на особенностях формирования нервных клеток, а также на эффектах отдельных генов в этих процессах. Очевидно, четко разграничить указанные группы вопросов удается далеко не всегда.
Также как и в биологии развития в целом, ключевой вопрос в изучении нейрогенеза — определение того, как (и почему) экспрессия группы генов происходит в данный момент и в соответствующем участке ткани. Понятно, что эти процессы регулируются не только генетической программой, действующей в конкретней группе клеток, но и внешними по отношению к нейронам процессами.
В качестве примера такого взаимодействия можно рассмотреть процесс нейрогенеза при доминантной неврологической мутации мыши Lurcher, при которой у гетерозиготных особей в возрасте 2 недель гибнут все клетки Пуркинье мозжечка. Эта гибель — результат прямого действия мутантного аллеля гена Lc именно в этих клетках. В то же время показано, что последующая дегенерация 90% гранулярных клеток — это вторичный эффект, следствие отсутствия клеток Пуркинье. Клетки Пуркинье в норме способствуют формированию нормальных синаптических контактов гранулярных клеток, а отсутствие нормально развитых синапсов ведет к постепенной дегенерации гранулярных клеток.
Очевидно, что нейрогенез представляет собой не серию сменяющих друг друга стадий, а процесс, в ходе которого происходит непрерывное взаимодействие сигналов, поступающих из внешней среды, и информации, считывающейся с генома. В схематической форме, однако, нельзя обойтись без условного деления процесса развития на стадии. Это деление проводится и в соответствии с современной эпигенетической концепцией. Последовательные стадии развития можно представить в виде схем, в которые входит ряд компонентов. Успех каждой стадии развития обеспечивается наличием следующих компонентов: фенотипа — Р, продуктов экспрессии определенных генов G, и существованием некоторого набора внешних условий, которые могут варьировать в определенных пределах:
Р1 + G1 + El -> Р2;
Р2 + G2 + Е2 -> P3 и т.д.,
где Р1 — это фенотип зиготы, Р2 — фенотип следующей стадии. При развитии нервной системы картина усложняется тем, что в категорию "внешних" условий попадают влияния, идущие от других, параллельно развивающихся глиальных и нервных клеток, которые по фенотипу и паттерну экспрессии генов не идентичны нейрону (или группе нейронов), интересующих нас в данный момент. Общий анализ показывает, что на нейрон действуют продукты экспрессии генов, которые можно условно разделить на 4 категории в зависимости от особенностей их экспрессии. Это гены, которые экспрессируются в дифференцирующихся нейронах, в нейронах других групп, в глиальных клетках и на уровне всего организма.
Схему хорошо иллюстрируют примеры развития нервной системы дрозофилы. Еще до появления первых эмбриональных закладок, во время образования трехслойного зародыша, на стадии нервного валика, т.е. в период, когда нервной системы еще нет, общий план ее строения уже начинает формироваться. Генетический контроль процесса в эти периоды трудно отделим от формирования общей схемы тела (подробнее см.; Корочкин, 1989, 1991, 1992).
Дрозофила. В настоящее время достаточно много известно о генах, контролирующих формирование общей схемы тела животного. Группы генов, экспрессия которых обеспечивает формирование общего плана строения эмбриона, обнаруживают неслучайный уровень гомологии у всех изученных организмов. Большая часть этой информации получена на дрозофиле.
Еще до оплодотворения, т.е в ооците, начинается экспрессия генов, продукты которых несколько позже принимают участие в формировании различий между передним и задним полюсом оплодотворенного яйца дрозофилы. Это так называемые "материнские" гены. К ним относятся, например, ген caudal (определяет полярность ооцита), ген bicoid (мутация по нему вызывает появление эмбрионов без головы и торакального ганглия), ген oskar (у мутантных эмбрионов нет брюшка).
Уже в самом эмбрионе проявляют свое действие гены других групп.
Больше всего информации получено по генам сегментации и по так называемым гомейозисным генам.
Гены сегментации. Мутации по ним нарушают метамерию, т.е. деление тела зародыша дрозофилы на сегменты, а также вызывают аномалии развития сегментов разных категорий. Могут происходить, например, потеря сегментов (ген gap), появление дефектных по структуре сегментов "через один" (ген pair rule), нарушение внутренней структуры полярности сегмента (engrailed). Совместное действие нормальных аллелей генов этих групп обеспечивает формирование сегментации тела. Мутации генов сегментации несовместимы с нормальным ходом эмбриогенеза и вызывают его остановку с последующей гибелью зародыша на разных стадиях.
Гомейозисные гены. Исторически первыми были описаны и изучены мутации по генам, которые вызывают заметные морфологические аномалии (уродства), но, тем не менее, совместимы с жизнью, т.е. не только не вызывают гибели зародыша, а даже не препятствуют метаморфозу и развитию имаго. Это так называемые гомейозисные мутации, выражающиеся в формировании сегментов с "неправильными" органами. Наиболее известны среди них — мутации aristapedia и antennaepedia, когда вместо аристы или антенны на голове мухи формируется конечность.
Гомеобокс. Молекулярно-биологические исследования гомейозисных мутаций выявили в соответствующих участках ДНК короткую консервативную последовательность оснований, которая была названа гомеодоменом, или гомеобоксом (Нох) (подробнее см.: Дондуа, 1997). Гомеодомен кодирует небольшого размера белковую молекулу, которая может связываться с ДНК. Эти данные позволили предположить, что гомеобокс кодирует регуляторный белок, способный включать и выключать экспрессию генов в определенные моменты развития. Структурные особенности ДНК Hох-доменов подтверждают предположение, что продукты этих генов могут связываться с ДНК и регулировать транскрипцию.
Итак, действие генов ряда групп проявляется либо в оплодотворенном яйце, либо позднее, на разных стадиях формирования самого эмбриона.
Мышь. Если у дрозофилы экспрессия генов Нох связана преимущественно с формированием сегментов и/или их внутренней структуры, то у других организмов родственные последовательности Нох имеют более разнообразные функции. Такие же как у дрозофилы гомеобоксные последовательности выделены в геноме шпорцевой лягушки, мыши и человека. Наиболее интенсивная экспрессия этих генов у позвоночных происходит в ЦНС, в почках и в зародышевых клетках.
У мыши описано и клонировано не менее 20 локусов, содержащих гомеобокс (Hох-локусы). У ряда из них определена нуклеотидная последовательность. Большая часть Hох-содержащих генов локализуется у мыши на хромосомах 2, 6, И и 15.
Общность генетических механизмов развития схемы тела насекомых и позвоночных подтверждается многочисленными случаями гомологии в строении соответствующих генов дрозофилы и позвоночных, например гена полярности сегментов wingless и онкогена мыши int-1 или гена achaete-scute с онкогеном с-mуc. Содержащие гомеобоксную последовательность гомейозисные гены групп engrailed и inverted дрозофилы имеют гомологов в геноме мыши, которые экспрессируются в эмбриогенезе при формировании нейроэктодермы. Продукты их транскрипции проявляются в полосках ткани, располагающихся в складках нервного гребня.
Содержащие гомеобокс гены дрозофилы и мыши имеют сходство в пространственно-временной картине экспрессии в процессе развития. Как и у двукрылых, в эмбрионе млекопитающих эти гены экспрессируются вдоль ростро-каудальной оси эмбриона. Следует отметить, что исследование функций этих генов у млекопитающих, точнее, у одного из наиболее изученных генетических объектов — мыши — достаточно сложно, поскольку соответствующие мутации неизвестны, тогда как у дрозофилы, например, анализ подобных мутаций — один из ключевых методов исследования.
Вместо анализа мутаций при работе с мышью предложен ряд другах методов изучения роли генома на ранних стадиях развития эмбриона. Один из наиболее продуктивных — введение в зародыш генетического материала, который представляет собой или измененный ген, имеющийся у хозяина, или делецию по определенному участку хромосомы (в этом случае используют специальные приемы, которые "заставляют" экспрессироваться новый ген). Можно также вводить ген, выделенный из генома другого вида. Примером последнего типа эксперимента служит работа американского ученого Р. Палъмитера (R. Palmiter) с сотрудниками. Они создали трансгенных мышей с геном, кодирующим гормон роста крысы под "транскрипционным контролем" так называемого промотора металлотенеина, последовательности, которая экспрессируется практически во всех тканях. Несколько мышей, полученных таким способом, отличались очень крупными размерами (см. также: 8.6.3.4). Интеграцию в геном хозяина и экспрессию нужной ДНК последовательности обеспечивают технически сложные приемы, когда интересующий исследователя участок ДНК "сшивается" с последовательностями, экспрессия которых вызывается особыми физиологическими нагрузками.
С помощью трансгенных животных можно изучать фенотипические эффекты изменения "дозы" такого гена. Подобные эксперименты позволили установить, что во всех отделах ЦНС "трансгенного" эмбриона мыши в ростро-каудальном направлении имеются продукты транскрипции всех 7 генов блока Нох, введенного в геном.
Гомеобокс — это только один тип консервативной последовательности из всех других типов, выявленных у организмов разного уровня развития. Описаны и семейства генов, которые содержат белковые домены, выделенные в связи с исследованием разных функций. Гены сегментации дрозофилы Kruppel и hunchback имеют белковый домен, гомологичный транскрипционному фактору III, выделенному у шпорцевой лягушки, так называемому "белку с цинковыми пальцами". Ген Нох мыши, продукт которого также представляет собой белок с "цинковыми пальцами", локализуется в протосегментах заднего мозга эмбриона. Его продукты показывают экспрессию в порядке "через один сегмент". Такой метамерный рисунок в экспрессии гена — четкое молекулярно-генетическое доказательство существования сегментации мозга позвоночных.
Таким образом, дальнейшая дифференцировка мозга на отделы, по крайней мере частично, определяется событиями, которые причинно связаны с экспрессией генов "схемы тела". Сходство ряда участков генома, таких как гомеобоксные последовательности, у организмов, занимающих разное положение на систематической лестнице, показывает, что это еще один путь сравнительных исследований структуры и функции мозга.
Белки, которые кодируются самим гомеобоксом или связанными с ним генетическими элементами, выполняют регуляторные функции. Это означает, что появление продуктов экспрессии таких генов, в свою очередь, вызывает экспрессию (или репрессию) целого ряда генов, кодирующих либо структурные белки, либо сигнальные молекулы, которые должны регулировать уже следующий каскад событий.
Даже беглое знакомство с подобными данными показывает, что в процессе развития выявляется некая иерархия управления, в которой гены каждого данного уровня передают инструкции "вниз" генам следующего уровня. Это в целом соответствует общей схеме эпигенетической концепции. Если в каком-то звене системы возникает мутация, то нормальный ход процесса развития нервной системы оказывается нарушенным. Очевидно, что чем "ближе" к началу нейрогенеза относится работа гена, претерпевшего мутацию, тем более сильный дефект вызывает такая мутация.
У человека и мыши, которые среди млекопитающих изучены генетически наиболее полно, описано много десятков мутаций, затрагивающих функцию нервной системы. Часть подобных случаев — это проявление обширного плейотропного эффекта генов, имеющих первичный дефект, изначально не связанный с нервной системой (см.: 8.2.2), другие же мутации влияют на формирование мозга более непосредственно. К их числу относятся так называемые неврологические мутации мыши, изучение которых дает информацию о роли тех или иных аллелей таких генов в нейрогенезе. У дрозофилы также выявлены подобные системы генов.
У дрозофилы описаны "кластеры" (т.е. функционально объединенные группы) генов, совместная работа которых связана с дифференцировкой основных зародышевых закладок по "нервному", или "эпидермальному" пути дифференцировки. Это так называемые "нейрогенные" и "антинейрогенные" локусы. Нормальные аллели первых осуществляют в процессе развития влияния, благодаря которым часть эмбриональных клеток начинает дифференцироваться как эпидермальные. Мутации по этим генам вызывают аномалию развития, которая практически несовместима ни с дальнейшим процессом развитая, ни с жизнью организма: у них происходит так называемая "нейрализация" развития. Такие эмбрионы представляют собой сверхразвитую нервную закладку, в которой практически нет или очень мало эпидермальных клеток (отсюда название — нейрогенные локусы).
Примером нейрогенного влияния мутации одного из таких генов является доминантная мутация Notch, описанная еще в 1940 г. В гомозиготном состоянии она летальна, но обнаруживает лишь небольшой морфологический дефект жилкования крыла в гетерозиготном состоянии. У гомозиготных по этой мутации эмбрионов дрозофилы увеличено число развивающихся из эктодермы нейробластов, а ткани других производных эктодермы, например кожных покровов, развиваться не могут. У таких эмбрионов формируется нервная система, у которой общий объем клеток, по крайней мере, в 3 раза больше, чем в норме. Мезодерма остается неразвитой. Эффекты подобных мутаций на ход процессов развития общей схемы тела и основных органов у разных организмов рассматриваются в обзорных статьях Л.И. Корочкина (1989, 1991, 1992), посвященных процессам генетического контроля развития. Гипернейрализованные, гомозиготные по этой мутации эмбрионы, которые через некоторое время должны погибнуть, были использованы Л.И.Корочкиым и С.В.Савельевым как материал для трансплантации в мозг земноводного — шпорцевой лягушки Xenopus laevis — с целью изучения свойств развития подобных гетеротрансплантатов (Корочкин, 1992).
Симметричную группу генов, "ведающих" формированием эмбриональных нервных клеток — нейробластов, составляют так называемые "антинейрогенные локусы. Мутации этих генов дают спектр аномалий, при которых также нарушается развитие нервной системы (например, мутация по комплексу Ac-Sc).
Пример мутации одного из таких генов (или генного кластера) дает мутация Delta, также летальная в гомозиготном состоянии и также вызывающая аномальное жилкование крыла в гетерозиготе. Гомозиготные по этой мутации эмбрионы обнаруживают чрезмерное развитие эпидермальной ткани при аномально малом числе нейробластов.
Различная "судьба", т.е. направление будущей дифференцировки эмбриональных клеток-предшественниц ЦНС детерминируется на ранних этапах эмбриогенеза — на стадии бластулы у дрозофилы и стадии нервного гребня у мыши. Прослеживая судьбу поколений клеток в процессе развития ЦНС млекопитающих, и в частности мыши, исследователи делают заключение о том, что генетически детерминированная судьба клетки сказывается не столько на окончательном положении в мозге, которое она займет как потомок клетки-родоначальницы, сколько на функциональных свойствах и количестве нейронов данного класса.
Генетические исследования развития в их традиционном варианте возможны лишь при наличии генетической изменчивости по тому или иному признаку, т.е. при выявлении и анализе эффектов соответствующих мутаций. Примером могут служить упоминавшиеся выше неврологические мутации дрозофилы — shaker, hyperkinetic (вызывающие судороги при впадении в эфирный наркоз), мутации cacophony, fruitless, stick (с нарушениями процесса спаривания), а также неврологические мутации мыши — reeler, weaver, quaking, waltzer и др.
Однако в случае очень многих мутаций экспрессия аномального гена вызывает гибель зародыша. В то же время иногда оказывается возможным реконструировать функцию нормального аллеля данного гена. Если бы можно было выделить мутации нескольких генов, экспрессирующихся на последовательных стадиях пренатального онтогенеза, это дало бы достаточно полную картину развития нервной системы. Наиболее вероятно, что такую информацию можно будет получить, изучая развитие дрозофилы, не только потому, что у нее выявлены нейрогенные и антинейрогенные локусы (см.: 8.5.3.1), но и потому, что у этого вида можно использовать чрезвычайно плодотворный метод генетических мозаиков.
Сходный методический подход у мышей обеспечил создание химерных и трансгенных животных.
Генетические мозаики. С помощью этого метода удается «проследить судьбу измененных мутацией клеток и тканей на всем протяжении эмбриогенеза.
Насекомые. Истинными генетическими мозаиками считаются животные, клетки которых происходят из одной зиготы, но несут, тем не менее, определенные генетические различия. Наиболее известным примером таких мозаиков являются гинандроморфные особи, описанные у паразитических ос Habrabracon juglandi П. Уайтингом. Гинандроморфы, т.е. особи, имеющие ткани и самца, и самки, очень редко (1 случай из 1000— 10000) выводятся из аномальных яиц с двумя ядрами, из которых бывает оплодотворено только одно. Женские ткани развиваются из диплоидного ядра, а мужские — из неоплодотворенного, гаплоидного. Если ткани, развившиеся из оплодотворенного яйца, имеют четкие морфологические отличия, например измененный цвет глаз или аномальное жилкование крыльев, то легко видеть, что тело гинандроморфной особи представляет собой мозаику из тканей самца и тканей самки (рис. 8.18). Наблюдая за поведением таких ос, Уайтинг показал, что насекомые ведут себя либо как самец, либо как самка в зависимости от того, как распределяются на теле участки "самцовых" и исамочьих" тканей (см.: Эрман, Парсонс, 1984).
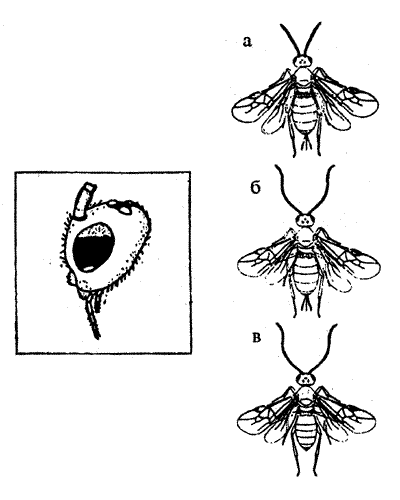
Рис. 8.18 Гинандроморфные особи Habrabracon juglandi. Для нормальной самки (а) характерны более длинные крылья, более короткие антенны и жало с парой сенсорных придатков, у самца (в) крылья короче, чем у самки, а антенны, длиннее, анатомия конца брюшка иная, б — гинандроморфная особь. Справа — голова гинандроморфной особи "в профиль" часть тканей глаза имеют генотип самца (светлые участки), а часть — генотип самки (темный участок)
У дрозофилы мозаичные по тканям самца и самки особи получаются, когда в скрещивание берутся самки с так называемой кольцевой Х-хромосомой. Эта кольцевая структура неустойчива и часто утрачивается в период ранних митотических делений эмбрионов женского пола, которые получили одну кольцевую, а одну нормальную Х-хромосомы. Клетка, в которой остается только одна Х-xpoмосома, дифференцируется по мужскому типу (поскольку у дрозофилы особи с генотипом Х0 — самцы). Мозаицизм обнаруживается по рецессивным генам, локализованным в Х-хромосоме, а также по тем признакам, развитие которых определяется генетической программой, свойственной одному или другому полу (рис. 8.19; 8.20).
Рис, 8.19. Схема получения гинандроморфных особей у дрозофилы (а); б — "карта судьбы": схематическое изображение бластулы дрозофилы, с указанием участков, в дальнейшем развивающихся в соответствующий орган и
Благодаря работам С. Бензера и его коллег, проведенным в 60—70-е годы, в настоящее время имеется достаточно полная картина так называемой "карты судьбы" ("fate map") — топографического расположения на бластуле участков, из которых впоследствии развивается тот или иной орган или часть тела дрозофилы (см.: Бензер, 1975), Основные успехи нейрогенетики и генетики поведения дрозофилы базируются именно на использовании метода гинандроморфных мозаиков.

Рис, 8.20. Схематическое изображение гинандроморфных особей Ткани самки — темные, самца — светлые, Разное соотношение тканей с разным генотипом связано со случайным процессом — утерей Х-хромосомы на разных стадиях дробления яйца
Млекопитающие. Единственный случай сходного типа мозаицизма у позвоночных описан еще в начале 70-х годов. Это исследование было посвящено мозаичности в проявлении мутации sh-1 мыши, затрагивающей строение кортиева органа. Анализировали фенотип самок мышей, гомозиготных по sh-1 и имеющих в гетерозиготном состоянии так называемую транслокацию Каттанаха (транслокация — это хромосомная перестройка, при которой фрагмент одной хромосомы соединяется с другой). При упомянутой транслокации фрагмент 7-й хромосомы с мутантным аллелем sh-1 оказался соединенным с Х-хромосомой. Как известно, в каждой из клеток тела самки в активном состоянии находится только одна Х-хромосома, а вторая инактиви-рована. Поскольку в случае данной транслокации ген sh-1 был соединен с одной из Х-хромосом, а мутация эта рецессивна, то в тех клетках, где была инактивирована Х-хромосома, несущая транслокацию (и ген sh-1), эффект мутантного гена не обнаруживался. Если же была инактивирована Х-хромосома, не имевшая транслокации, то оба гена sh-1 были активны и проявлялся эффект рецессивной мутации. Фенотипически это выражалось в том, что в кортиевом органе участки мутантных и нормальных клеток перемежались.
Химеры млекопитающих. Животных с генетически неоднородным организмом можно получать и в специально спланированных эмбриологичеких экспериментах (Мак-Ларен, 1979) — речь идет о химерах. Получение химер млекопитающих состоит из нескольких этапов. При использовании агрегационного метода извлеченные из половых путей самки эмбрионы на стадии дробления приходят в соприкосновение друг с другом, в результате чего они сливаются в один. С помощью инъекционного метода в бластоцисту эмбриона вводятся клетки другого эмбриона или другие донорские клетки (например, клетки определенной опухоли, клетки костного мозга и т.д.). На следующем этапе химерный эмбрион переносят в матку приемной матери.
Для распознавания отдельных компонентов химерного организма необходимо, чтобы ткани разного происхождения имели четкие и простые маркеры. Таким маркером может быть особенность пигментации, наличие редких форм изозимов, антигены, хромосомные транслокации и т.п.
Получение химерных животных в настоящее время налажено во многих лабораториях. Проблема подбора адекватных маркеров (морфологических — цвет шерсти, или биохимических — изоферменты) для анализа "происхождения" того или иного участка мозга химерного животного, как правило, бывает достаточно трудной. Для такого маркирования используются, например, межлинейные генетические различия в локусе структурного гена бета-галактозидазы. Этот фермент обнаруживается цитохимически в крупных нейронах. Нередко используют также антигенные различия в генопродуктах разных аллелей гена глюкозофосфатизомеразы.
Большинство неврологических мутаций мыши (это, как правило, аномалии строения мозжечка и/или внутреннего уха) рецессивны, т.е. их эффект проявляется в гомозиготном состоянии, а гетерозиготные особи практически не отличаются от нормы. В то же время особи, гомозиготные по этим мутациям, имеют резко сниженную жизнеспособность и поэтому трудны для изучения. Получение мышей-химер, составленных из нормальных и мутантных тканей, позволяет изучать мозг, мозаичный по генетическому составу. При удачном подборе маркеров можно увидеть, что даже рядом расположенные нейроны одного класса (например, клетки Пуркинье) имеют разный генотип. Как правило, неврологический дефект у таких химерных мышей не проявляется, однако на нейроанатомическом уровне можно установить, какой именно тип клеток несет первичный дефект мутации, а какие клетки страдают вследствие нарушений нормальных эпигенетических отношений с первично аномальными. Данные по мутации Lurcher, упоминавшиеся выше (см.: 8.5.1), были получены именно таким методом. Анализ особенностей поведения мышей-химер также представляет интерес для выяснения роли генотипа в определении особенностей той или иной реакции. С мутацией Lurcher, например, были проведены эксперименты, показавшие, что у гетерозигот LC1+ сильно нарушена локомоция, тогда как у мышей-химер, "составленных" из эмбрионов, гомозиготных по этой мутации, и нормальных зародышей, таких изменений не было (см. также: Goldowitz et al., 1992).
На серийных срезах мозга, например на срезах ствола и мозжечка химерных мышей, у которых нормальные и мутантные нейроны различаются визуально, можно определить общее количество нейронов в моторном ядре лицевого нерва, а также число клеток Пуркинье мозжечка с нормальным и мутантным фенотипами. Статистические оценки позволили заключить, что родоначальниками клеток Пуркинье в мозге мыши являются 8 клеток, а нейроны ядра лицевого нерва происходят от 12 клеток-предшественниц.
У мышей известны межлинейные различия по числу нейронов той или иной структуры мозга. Есть веские основания предполагать, что одной из причин этой изменчивости могут быть различия в числе клонов, формирующих данный участок мозга, т.е. в числе "клеток-родоначальниц" и/или в размере клона, т.е. в числе митотических делений исходной клетки перед началом дифференцировки.
Нейрохимические, морфологические и физиологические различия между инбредными линиями мышей, описанные к настоящему времени, несомненно, определяются значительным числом локусов. Современные методы генетического анализа, использующие множественные маркеры, а также конгенные и рекомбинантные линии (в частности, метод картирования QTL — quantitative trait loci), позволяют описывать такие межлинейные различия. Возможно, они помогут и в описании структурных различий в мозге, которые являются ключевыми в возникновении межлинейных различий в поведении.
Трансгенные мыши. На основе метода получения химерных животных был разработан принципиально новый подход к исследованию генетических закономерностей формирования организма животного. Он получил название "метода создания трансгенных животных", о котором говорилось в разделе 8.5.2. Этот метод позволял исследовать особенности развития мозга и поведения животных, у которых были искусственно изменены или "выбиты" (отсюда название "нокаут", knock-out) ген или гены, кодирующие определенные структурные и/или регуляторные белки (см.: 8.6.3.3 и 8.6.4.3).
Используя мутации, затрагивающие морфогенез мозга, прижизненное окрашивание отдельных клеток с последующим прослеживанием распределения метки, а также создание мышей-химер, можно проследить судьбу отдельных клеточных клонов при развитии мозга и выявить характер взаимодействия между ними. Важным свойством дифференцировки нейробластов млекопитающих является определение окончательных свойств нейронов на таких стадиях развития, когда фенотипически клетка еще не дифференцирована.
Какие же характерные для нейрона свойства должна проявить такая рано детерминированная клетка в ходе дифференцировки? К ним относятся: особенности морфологического строения, медиаторная специфичность, определенный характер ее связей с другими нейронами, общее число нейронов данного фенотипа, а также определенный численный баланс клеток разных популяций.
Очевидно, что генетическая программа развития нейрона развертывается на основе тех эпигенетических механизмов, о которых упоминалось выше. Все перечисленные выше свойства детерминированной нервной клетки формируются как результат экспрессии со ответствующих генов в определенные моменты времени, которые совпадают с формированием (вследствие работы других групп генов) градиентов концентрации биологически активных веществ (гормонов, факторов роста и др.) — компонентов внешней среды нейрона, необходимых для его развития.
Большой фактический материал по развитию ЦНС, в частности, при неврологических мутациях мыши, заставляет сделать вывод, что мутационные события или внешние экспериментальные воздействия могут значительно повлиять на окончательное пространственное положение нейронов, но не на их форму или функциональные свойства. Ниже это иллюстрируется более подробным описанием неврологических нарушений при некоторых мутациях.
Следует отметить, что изучение неврологических мутаций мыши дало огромный материал для понимания генетического контроля пре- и постнатального онтогенеза мозжечка, Особое значение, которое придается этим данным, связано с тем, что подобные мутации могут служить удобными и полезными моделями генетических дегенеративных заболеваний человека. Это относится прежде всего к мутациям "демиелинизации", например jimpy, shiverer и др. Молекулярная биология этих мутантных генов изучена достаточно подробно.
У собак описана мутация демиелинизации — "shaking pup". Она рецессивна и сцеплена с Х-хромосомой. Дефекты этого типа затрагивают синтез эволюционно консервативных белков миелина. Демиелинизация, при которой ухудшаются условия проведения нервных импульсов по аксонам, сопровождается снижением всех функций мозга и гибелью животного. При "shaking pup" дефект, по всей видимости, связан с заменой одной пары оснований в гене, кодирующем один из таких белков. Столь сильные изменения в функциональных свойствах белков миелина определяются, по всей видимости, именно большим эволюционным консерватизмом белков этой группы. У мутантных особей, начиная с 10—12-дневного возраста, появляется мышечный тремор (характерный для мутаций демиелинизации у всех видов млекопитающих). У мутантных собак, в отличие, например, от некоторых мутаций мышей, миелин разрушается не полностью, и они живут относительно дольше, чем мутантные крысы и мыши. В то же время у 4—16-месячных мутантных щенков количество миелина составляет лишь 2,4% от контроля. У таких собак резко снижено количество олигодендроцитов — глиальных клеток, продуцирующих миелин.
Неврологические мутации мыши имеют аутосомную локализацию. Они, за редкими исключениями, рецессивны, их наиболее частый и наиболее заметный морфологический дефект — недоразвитие органов внутреннего уха и/или мозжечка. Мыши с неврологическими мутациями характеризуются нарушениями движений, походки, аномалиями позы, отклонениями в поведении и в способности к обучению.
Нейроны большинства корковых структур мозга млекопитающих образуются в генеративных зонах, которые пространственно отделены от мест расположения этих клеток во взрослом мозге. Так, предшественники нейронов древней (обонятельной) и старой (аммонов рог) коры, а также новой коры формируются в вентрикулярных и субвентрикулярных зонах переднего мозга. Предшественники гранулярных клеток зубчатой фасции гиппокампа формируются in situ, предшественники клеток Пуркинье — в вентральной зоне вдоль дорсального края 4-го желудочка, а разные классы клеток коры мозжечка образуются в пространстве между наружным гранулярным слоем и развивающимся молекулярным слоем.
По окончании митозов большая часть корковых нейробластов мигрирует на значительные расстояния. На окончательных позициях они распределяются по слоям (выявляется дифференцировка на классы) в направлении (в случае корковых структур) от внутреннего слоя к наружному.
Изучение мутаций reeler и weaver показало, что на окончательное положение нейронов в слоях влияют два независимых нейрогенетических механизма — миграция клеток и распределение их по слоям. В то же время ключевые события, вызывающие нарушения и гибель клеток наружного гранулярного слоя мозжечка, происходят еще раньше (примерно в первый день после рождения).
При мутации reeler развитие гранулярных клеток замедляется, отмечается также их ускоренная гибель. Нарушение цитоархитектоники мозжечка, а в случае мутации reeler еще коры и гиппокампа, — лишь наиболее заметный эффект этой группы неврологических мутаций (рис. 8.21).
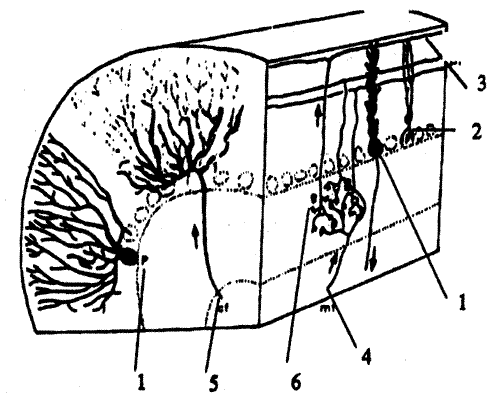
Рис. 8.21. Упрощенная схема синаптических контактов основных групп клеточных элементов мозжечка. 1- клетки Пуркинье; 2 — бергмановская глия; 3 — параллельные волокна; 4 — мшистые волокна; 5 — лазящие волокна; 6 — гранулярные клетки
У reeler выявлены отклонения в строении ряда ядер продолговатого мозга и базальных ганглиев, а также в особенностях распределения на нейронах рецепторов разных классов. При этой мутации (rl) нарушено расположение слоев корковых структур. Аномальная стратификация обнаруживается у них с момента закладки корковой пластинки (13—14-й дни эмбрионального развития). Полиморфный слой клеток располагается на поверхности, слои мелких пирамид (в норме это 3-й и 4-й слои) находятся в глубине, тогда как 5-й слой нормального мозга — ближе к поверхности. Характер распределения восходящих моноаминергических волокон в подкорковых структурах не отличается от нормы, однако их распределение по слоям коры у rl имеет характерные черты. Эти афференты не ветвятся в наружном слое коры и не идут каждое отдельно, а собираются в пучки. В тоже время места их окончаний в нормальном и мутантом мозге гомологичны.
Таким образом, нарушение архитектоники слоев новой коры не препятствует "узнаванию" растущими волокнами соответствующих клеток-мишеней. На срезах мозга мышей, гомозиготных по rl, было показано, что при этой мутации ослаблен процесс формирования долговременной постсинаптической потенциации, а в stratum radiatum и stratum moleculare-lacunosum присутствует множество клеток, иммунореактивных к парвальбумину (нередко это свойство находят у ГАМК-ергических вставочных нейронов).
Нормальный аллель гена rl обеспечивает, таким образом, сопряжение во времени и в пространстве процессов миграции и созревания клеточных элементов.
Считается, что наиболее правдоподобны два объяснения причин дефекта строения мозга при мутации rl: уменьшение сил сцепления (адгезии) между нейронами и волокнами радиальной глии к концу периода миграции и изменение этих сил в постмиграционный период. Было действительно продемонстрировано, что в коре мышей-мутантов по rl глиальные волокна располагаются тангенциально к боковым стенкам формирующихся "бочонков" — специфических агрегаций корковых нейронов, а не радиально, как в норме. Полагают, что в развивающейся коре радиальные глиальные волокна служат "гидами" для мигрирующих молодых нейробластов, которые проделывают свой путь к коре из вентральной зоны, где происходят митозы.
При исследовании гистогенеза мозжечка у мышей-химер, т.е. у животных, ткань мозга которых состояла из нормальных и мутантных клеточных элементов, была подтверждена роль глиальных элементов. В целом для мутации rl характерно общее недоразвитие мозжечка с почти полным отсутствием клеток-зерен. Клетки Пуркинье у этих мутантов (а также при ряде других мутаций) развиваются в отсутствие контактов с аксонами гранулярных клеток — параллельными волокнами, вместо которых с клетками Пуркинье контактируют мшистые волокна, в норме приходящие в гранулярный слой. При этой мутации клетки Пуркинье расположены беспорядочно и не формируют четкого слоя. У мышей-химер аномальные участки мозжечковой коры "вкраплены" в кору с нормальной структурой. В то же время клетки Пуркинье, которые маркированы как генотип rl/rl, можно найти как в нормальных, так и в мутантных участках ткани.
Согласно результатам иммуногистохимического исследования мозжечка, влияние мутации rl на миграцию элементов может осуществляться в более ранние сроки, еще до начала миграции. Возможно, что эффект действия мутантного гена начинается в период формирования нейробластов.
У мышей-химер, "составленных" из нормальных клеток и генотипа staggerer (sg), в клетках Пуркинье мутантного фенотипа присутствуют все аномалии строения, свойственные этой мутации. Известно, что у мутантов sg гранулярные клетки мигрируют и дифференцируются нормально, но к 4-й неделе жизни начинают прогрессивно дегенерировать, а клетки Пуркинье при этой мутации сохраняются в количестве 10% от нормы.
При мутации weaver (wv) дефект обусловлен отсутствием миграции гранулярных клеток и последующей их дегенерацией. Таким образом, мутантный локус действует непосредственно на гранулярные клетки.
Примером еще одного типа нарушений регуляторных процессов нейрогенеза является мутация stambler, при которой структура мозжечка не дезорганизована, но общее число и гранулярных клеток, и клеток Пуркинье уменьшено. В определенный период развития в этих нейронах начинаются нарушения внутренней структуры, которые, однако, затем исчезают.
Приведенные примеры показывают, что метод создания мозаичных особей и метод создания химерных мышей, позволяют исследовать функции генов, реализующих эффекты на стадиях органогенеза.
В процессах развития нервной ткани важная роль принадлежит временному режиму экспрессии генов, механизмы которого тесно связаны с процессами межтканевых и межклеточных взаимодействий.
Экспрессия специфических генов в определенные моменты времени определяет, каким будет соотношение медиаторных или гормональных продуктов в конкретный период развития, и как будут формироваться морфологические особенности мозга — его отдельные структуры и связи между ними.
Рассмотрим некоторые особенности формирования медиаторной специфичности гипоталамуса. В одной из первых работ в этой области было показано, например, что в паравентрикулярных и супраоптических ядрах гипоталамуса процессинг (т.е. формирование малых пептидных молекул из белков-предшественников с помощью ферментов-протеаз), обеспечивающий "наработку" окситоцина и аргинин-вазопрессина, происходит в разные сроки онтогенеза. В результате этого, начиная с определенного момента развития, нейроны названных ядер имеют четкие нейрохимические различия.
Особенности экспрессии генов в мозге исследуются в настоящее время все более интенсивно. Такие методы позволяют, с одной стороны, оценивать функциональную роль тех или иных воздействий на мозг, точнее, на его генетический аппарат. С другой стороны, нарастает число работ, в которых исследуются изменения экспрессии специфичного для нервной ткани гена при каких-либо воздействиях, например при сенсорной стимуляции, при стрессе, разрушении отделов мозга и т.п. В этом отношении можно уверенно утверждать, что рождается новый методический подход к оценке физиологического состояния мозга, сходный с методом исследования электрофизиологических процессов в нервной ткани. Наиболее полно в этром плане изучены так называемые протоонкогены, или быстрые гены раннего действия, относящиеся к семействам генов c-fos, c-jun и др.
Эти гены кодируют белки, являющиеся так называемыми факторами транскрипции, т.е. молекулами, которые определяют включение в работу отдельных генов и групп генов, или же их репрессию. Не вдаваясь в детали описания этих молекулярных событий, отметим, что гены раннего действия были первоначально описаны как факторы, способствующие или сопутствующие формированию злокачественных новообразований. Позднее было показано, что в нормальных условиях эти гены интенсивно экспрессируются в период эмбрионального и частично постэмбрионального развития. В то же время их экспрессия проявляется через очень короткий промежуток времени после некоторых физиологических, а также патологических воздействий на мозг. Примером может служить повышение экспрессии гена c-fos как реакция на развитие судорожной активности. Поскольку экспрессия этих генов оказывается, видимо, первым событием в каскаде, ведущим к изменению экспрессии генетических элементов в ряде структур мозга, то очевиден большой интерес исследователей к ним (см.; Анохин, Судаков, 1993). У крыс линии КМ, выделенной по высокой чувствительности к звуку (см.: 8.6.5.1), в период после судорожного припадка экспрессия генов этой категории оказывается сниженной.
Иммуногистохимическим методом в ткани мозга крыс выявлены специфические белковые продукты экспрессии ранних генов. Они были найдены в нейронах примерно 100 ядер и областей мозга интактной крысы (c-Jun JunB, JunD, c-Fos, FosB, Krox-24). Был составлен своеобразный "атлас" экспрессии соответствующих ранних генов (Herdegen et al., 1995), каждый из которых имел свой "индивидуальный профиль" экспрессии в мозге. Полагают, что активация семейств таких генов определяется разными механизмами и происходит при разных физиологических состояниях мозга и организма в целом.
Классический менделевский анализ рассматривает признаки, которые находятся под влиянием одного, двух, реже трех генов. Это так называемое моно- или дигенное (олигогенное) наследование. В подобных случаях фенотипы разделяются на небольшое число дискретных классов. К категории "менделирующих" генов относятся, например, гены, влияющие на обмен веществ, а их плейотропные эффекты затрагивают поведение (см.: 8.2.2). Ряд примеров такого наследования особенностей поведения, например уровня двигательной активности, будет приведен в разделе 8.6.4.
В то же время огромное большинство признаков поведения, так называемых количественных, отличаются плавной (недискретной) изменчивостью, которая определяется значительным числом генов. В генетике количественных признаков анализируют состав изменчивости. Ее оценивают путем определения генетической и средовой компонент дисперсии величины признака, а также доли изменчивости, которая связана с эффектами взаимодействия генетических и средовых факторов. Эти методы подробно рассматриваются в специальных руководствах (Рокицкий, 1977; Мазер, Джинкс, 1985 и др.). В применении к генетическим исследованиям поведения примеры таких расчетов и пояснения к ним можно найти в руководстве Эрман и Парсонса (1984).
Ниже рассматриваются основные результаты генетических исследований как олигогенных, так и количественных признаков поведения, а также некоторые данные, которые легли в основу современного понимания роли генотипа в процессах формирования поведения. Кроме того, дается краткое описание основных методов и экспериментальных приемов.
Несмотря на то что основные результаты в генетике поведения получены на лабораторных грызунах, важным объектом исследования была и собака. К изучению генетических основ поведения этих животных предрасполагало их крайнее разнообразие по экстерьеру и по поведенческим реакциям.
Собака была традиционным лабораторным животным в исследованиях И.П.Павлова. Именно для собак была впервые сформулирована его концепция типов высшей нервной деятельности и выделены 4 основных типа (или темперамента), условно названных холерическим, сангвиническим, флегматическим и меланхолическим, а также некоторое число промежуточных по характеру подтипов. Каждый из этих типов ВНД предполагал у его носителя определенную комбинацию более простых свойств нервной системы — возбуждения и торможения, т.е. силы, подвижности и уравновешенности. Типология ВНД, предложенная Павловым, отражала стремление подвести строгую научную основу и дать количественную оценку индивидуальной изменчивости признаков поведения.
Следующим логическим этапом такого подхода было изучение генетических основ индивидуальной изменчивости ВНД, точнее, генетических основ типологии. Однако проведение генетических исследований оказалось малосовместимым с длительной и очень трудоемкой процедурой определения типа ВНД, принятой в лаборатории Павлова. В том виде, в каком исследования были задуманы, они оказались невыполнимыми. В то же время было изучено наследование более простых характеристик условнорефлекторной деятельности у собак, в частности проанализировано наследование силы нервных процессов (Крушинский, 1991).
Наиболее простым методически и в то же время весьма перспективным в плане изучения генетико-физиологических механизмов поведения оказался анализ наследования общего уровня возбудимости животного.
Переход к оценке возбудимости как одного из базовых свойств, обеспечивающих проявление и выраженность генетически детерминированных особенностей поведения собак, послужил далее основой для фундаментальных классических исследований генетики поведения этих животных, проведенных Л.В.Крушинским. Результаты этих исследований, осуществленных в 30—40-е годы, были изложены в ряде оригинальных статей тех лет, а также в первой монографии Крушинского (1962). В настоящее время эти работы можно найти в первом томе двухтомного издания избранных трудов Л.В. Крушинского (1991).
Первым этапом исследований Крушинского было подыскание адекватного и достаточно надежного метода оценки уровня общей возбудимости животного. В это понятие входит представление не только о некоем фоновом уровне возбудимости ЦНС, при котором протекает вся деятельность организма и, в частности, его поведение, но и о реактивности ЦНС к внешним воздействиям. Однако по мере проведения исследований автор пришел к выводу, что адекватным и достаточно надежным показателем общей возбудимости ЦНС собаки может служить уровень ее двигательной активности. Этот признак можно было просто и быстро оценивать с помощью спортивного шагомера, крепившегося на теле.
Основным генетическим экспериментом этой работы был следующий. Гиляцких лаек, маловозбудимых собак, не обладавших пассивно-оборонительной реакцией (т.е. не трусливых), скрещивали со значительно более возбудимыми немецкими овчарками, у которых пассивно-оборонительная реакция также отсутствовала. У потомства первого поколения (25 особей) была резко выражена пассивно-оборонительная реакция. Подобное проявление этой реакции в каком-то смысле не было неожиданным, поскольку в одном из более ранних исследований такое проявление трусости было обнаружено у гибридов Fl немецких овчарок с волками. В случае волко-собачьих гибридов их выраженная пассивно-оборонительная реакция, также не отмеченная у родителей, была приписана специфическим особенностям генотипа волка. В случае скрещивания лаек и овчарок дикие предки в родословной отсутствовали, и причиной повышенной трусливости потомства был какой-то иной фактор. В то же время характерной особенностью и волков, и гиляцких лаек была достаточно низкая общая возбудимость, проявлявшаяся в невысоком уровне двигательной активности обеих групп животных.
Л.В. Крушинский предположил, что у гиляцких лаек существует породная предрасположенность к проявлению пассивно-оборонительной реакции, но она не обнаруживается у них из-за низкого уровня возбудимости. Гибриды этих собак с овчарками унаследовали от последних высокую возбудимость, а от другого родителя — пассивно-оборонительную реакцию, в результате чего это свойство поведения — повышенная трусливость — было выражено у них в полной мере. Такое объяснение было подтверждено простым экспериментом с введением гиляцким лайкам кокаина, дофаминергического агента, который специфически активировал двигательную сферу животного. У гиляцких лаек (но не у собак других пород), получавших инъекции кокаина, четко проявлялась пассивно-оборонительная реакция. Таким образом, уровень возбудимости, который в данных экспериментах коррелировал с уровнем двигательной активности, оказался модулятором проявления других признаков поведения.
Таким образом, генетически детерминированный высокий уровень признака поведения может не обнаруживаться у животных с маловозбудимым фенотипом. В то же время у потомства, полученного от скрещивания таких собак с возбудимыми особями, подобный признак был достаточно отчетливым.
В середине 40-х годов было начато наиболее полное и детальное исследование генетики поведения собак. Результаты этой многолетней работы были изложены в монографии Дж. Скотта и Дж. Фуллера (Scott, Fuller, 1965), которая увидела свет в 1965 году, но продолжает оставаться одной из наиболее серьезных книг на эту тему. Объектами исследования были собаки 5 пород — бассенджи, бигли, американские коккер-спаниели, шетландские овчарки (шелти) и жесткошерстные фокстерьеры, а также гибридные животные. Помимо сравнения поведения собак разных пород и их гибридов, авторы были заинтересованы в изучении формирования поведения собаки в онтогенезе. Для этого они проводили скрупулезные систематические наблюдения щенков, начиная от рождения и до возраста 16 недель. Всего было исследовано несколько сотен животных.
Первый этап работы заключался в подборе пород собак и методик тестирования поведения. При исследовании развития поведения в ряде случаев щенков воспитывали "перекрестно", т.е. часть помета выращивалась биологической матерью, а часть — приемной, причем другой породы. Небольшое число собак всех пород было выращено не в питомнике, а в семьях. Если поведение выращенных в домашних условиях собак по отношению к человеку и было несколько иным, чем собак питомника, то показатели большинства тестов практически не отличались от тестов у остальных животных. Специальное внимание исследователи уделяли условиям содержания, выращивания и кормления животных, стремясь обеспечить их максимальное единообразие.
На втором этапе исследования проводился генетический эксперимент, для которого были выбраны две заведомо неродственные между собой породы собак — бассенджи и американские коккер-спаниели. Бассенджи — это неспециализированная порода охотничьих собак некоторых африканских племен. Они отчетливо агрессивны и по ряду признаков напоминают диких животных. Коккер-спаниели — напротив, порода с низкой агрессивностью и достаточно долгой историей разведения.
По данным эксперимента, относительная дикость бассенджи проявляется у щенков при попытках взять их в руки. Они ведут себя подобно детенышам диких животных: издают резкий визг, пытаются вырваться, укусить и т.п. Кроме того, они резко сопротивляются попыткам приучить их к поводку. Гибриды первого поколения сходны по этим признакам с бассенджи. Картина расщепления признаков у животных второго поколения и у бэккроссов показала, что "дикость" бассенджи контролируется одним доминантным геном. Сопротивление ограничению свободы, по-видимому, также контролируется одним геном, но без доминирования. В этом случае нельзя исключить и материнское влияние.
Авторы проанализировали и такой признак, как агрессивность в игре, которая, по их данным, может контролироваться двумя генами без доминирования, но может иметь и более сложное определение.
Голосовые реакции авторы анализировали у щенков разного возраста в тесте на доминирование, когда двух животных заставляли бороться за обладание костью в течение 10 минут. Голосовые реакции имели два четких и независимых признака. Один из них — это порог провокации лая, т.е. уровень стимуляции, необходимый для проявления реакции. Он оказался высоким для бассенджи и низким — для коккер-спаниелей. Второй признак — длительность лая. Она была малой у бассенджи и большой у коккер-спаниелей. По данным скрещиваний, низкий порог проявления лая доминирует над высоким, а частота и длительность лая наследуются по промежуточному типу. Авторы полагают, что наследование уровня порога провокации определяется двумя независимыми доминантными генами, тогда как длительность лая не доминирует ни по высоким, ни по низким значениям и зависит, по-видимому, от одного гена.
Подобным образом были проанализированы и другие признаки поведения, наследование которых также оказалось под контролем одного или двух генов. Представляет, в частности, интерес наследование характеристики эстрального цикла самок. У бассенджи течка наступает один раз в год и строго приурочена к осеннему сезону. У коккер-спаниелей она бывает 1 раз в 6 месяцев в любой из сезонов года. Более подробно результаты этой работы описаны в монографии Эрман и Парсонса (1984).
Скотт и Фуллер оценили результаты своих исследований как довольно неожиданные, поскольку для ряда достаточно сложных признаков поведения было продемонстрировано моно- и олигогенное наследование. Они отметили, что взятые для исследования породы собак были в достаточной степени контрастны по поведению, видимо, вследствие разного давления отбора при их формировании. Был сделан вывод, что породное разведение собак, хотя и не создает полной инбредности, но способствует появлению гомозиготности по многим аллелям. Если в скрещивании участвуют две неродственные между собой породы (как было в данном случае), то у второго поколения гибридов можно получить картину расщепления, которая свидетельствует о контроле величины признака одним или двумя генами.
Именно здесь уместно вспомнить те рассуждения, которые приводились выше (см.: 8.2.5). Речь шла о том, что при оценке внутривидовых вариаций поведения практически всегда обнаруживается изменчивость по частоте выполнения той или иной поведенческой реакции или движения (ФКД), тогда как форма таких движений бывает неизменной. Крайним случаем вариации частоты ФКД может быть его крайне редкое проявление в репертуаре поведения собак данной породы, т.е. практически почти полное отсутствие. Если изменчивость порогов реакций и частоты их выполнения достаточно велика, то всегда можно подобрать такие внешние условия, которые могут спровоцировать появление реакции, даже если порог ее проявления в нормальных условиях достаточно высок.
С самого начала Скотт и Фуллер отметили, что у собак исследованных пород величины практически всех признаков в разной степени перекрывались, т.е. не было признака, который наблюдался бы только у одной породы и полностью отсутствовал бы у других. Авторы сделали вывод, что не существует какого-либо "породного набора признаков", поскольку в любой породе можно найти особей, чьи специфические признаки поведения перекрываются признаками другой породы. По данным их работы, олигогенное наследование характерно только для некоторых признаков поведения, а во многих случаях распределение признаков у потомства свидетельствовало о полигенном наследовании. Это в первую очередь относится к поведению собак при сложных тестах, включающих обучение в лабиринте, дифференцировку и тест на оперирование пространственными представлениями.
Помимо установления сложного полигенного наследования способностей к решению сложных тестов было показано, что при определении успешности решетая теста собаками разных пород важную роль играют и "некогнитивные" переменные. В частности, межпородные различия в успешности решения задач могли определяться разным уровнем трусливости, тогда как истинные градации по уровню собственно когнитивных способностей выявлялись с трудом.
Это обширное исследование, проведенное в начальный период развития генетики поведения и на таком сложном объекте, как собака, тем не менее, предвосхитило основные результаты, полученные в этой области в дальнейшем. Позднейшие генетические исследования сложных форм поведения лабораторных грызунов выявили важную роль в их осуществлении более простых переменных (см., например: Эрман, Парсонс, 1985).
Первыми исследованиями по генетике поведения были попытки селекции крыс на способность к обучению. Большинство из этих работ имеет лишь исторический интерес, однако проведенная С. Трайоном работа по отбору крыс на быструю и медленную обучаемость в сложном лабиринте при пищевом подкреплении составляет исключение. Начатая в середине 20-х годов, она увенчалась созданием 2 линий, получивших название Trayon maze-bright (TMB) и Trayon maze-dull (TMD), которые существуют по сей день и поддерживаются без дополнительного отбора (см. рис. 8.22).
В многочисленных исследованиях способности к обучению и ряда физиологических признаков у крыс этих линий (см.: 8.6.3.1) было показано, что успешность обучения в лабиринте "умной" линии и большое число ошибок у "глупой" специфичны только для лабиринтов данного типа, а при других тестах на обучение результаты могут иметь даже обратный знак.
Выведение линий, различающихся по каким-то параметрам поведения, представляет собой обычно лишь первый этап работы. Если такие линии различаются не только по величине исследуемого признака, но и по другим параметрам, то подобную ассоциацию необходимо изучить в специальных экспериментах.
Искусственный отбор проводится на базе исходной популяции животных, которая всегда бывает довольно малочисленна. При малом числе животных-основателей линии всегда существует достаточно большая вероятность случайной "фиксации" у отбираемых по нашему признаку аллелей других генов, не имеющих отношения к изучаемому признаку поведения. Поэтому корреляцию каких-либо двух и более признаков у селектированных линий нельзя трактовать как свидетельство общности их физиологической основы. Это одно из основных правил в исследованиях по генетике сложных форм поведения. Чтобы доказать наличие общих причин изменчивости таких коррелированных признаков, необходимо провести дополнительные исследования.
Проиллюстрируем это на следующем примере. В работе голландского ученого X.Ван-Абилена на основе популяции гибридов между линиями мышей C57BL/6J и DBA/2J были выведены две линии, контрастные по уровню исследовательской активности в установке ''открытое поле", точнее по количеству исследовательских "стоек".
Данные линии были также контрастны и по уровню активности ацетилхолинестеразы в коре головного мозга. Однако у гибридов второго поколения и у беккроссов этой корреляции не было, что свидетельствует об отсутствии непосредственной связи между признаками. В то же время (см.: следующий раздел) была обнаружена связь исследовательской активности с показателями некоторых мозговых медиаторных систем, но она имела значительно более сложный характер.
Если два признака продолжают обнаруживаться в тесной, статистически достоверной ассоциации друг с другом у гибридов F2 и У беккроссов, то предположение об их тесном сцеплении бывает оправданным. Примером логично построенной работы по доказательству связи способности к обучению реакции активного избегания в челночной камере и размером интра- и супрапирамидальных полей окончаний мшистых волокон в поле САЗ гиппокампа является серия исследований Х.-П.Липпа и др. (см.: 8.6.7.2).
Другой способ оценки случайности или неслучайности корреляции признаков поведения при селекции — это проведение двух (или более) параллельных селекционных экспериментов. В этом случае случайная фиксация аллелей, если она произошла в одной из пар селектируемых линий, с очень высокой вероятностью не произойдет в другой паре, и вопрос о генетической природе обнаруженной корреляции будет выяснен. Так, например, при селекции мышей на большой и малый вес мозга, проведенной в четырех независимых экспериментах (см.: Wimer et al., 1969; Fuller, 1979; Попова, Полетаева, 1986; Попова и др., 1977), было обнаружено, что мыши с большим весом мозга обладают достоверно более высокой обучаемостью навыкам разной природы. Это означает, что действительно больший вес мозга является фактором, благоприятствующим обучению. Отметим, что на человека эта закономерность не распространяется.
Исследователи уделяют большое внимание поиску физиологических механизмов межлинейных различий в поведении. В этом отношении интересны работы, проводимые в Институте цитологии и генетики РАН в Новосибирске (Трут, 1987). Начатые много лет назад длительные селекционные эксперименты по выведению линий (лисиц, норок, крыс-пасюков), различающихся по агрессивности, увенчались успехом, и исследователи имеют в своем распоряжении уникальные "параллельные" селектированные линии представителей разных отрядов млекопитающих (об этологических представлениях поведения при конфликте см.: 4.3).
Поскольку физиологическая регуляция признака "агрессия по отношению к человеку" изучена достаточно хорошо, поиск генетических коррелятов повышенной и пониженной агрессивности производился не вслепую, а на основе уже имевшихся представлений о ключевой роли биогенных аминов и системы их синтеза и деградации в определении уровня агрессивности. Очевидно, что и в таких случаях важную информацию может дать анализ корреляции признака в расщепляющихся поколениях. Однако сам признак хорошо изучен, и именно благодаря этому для выяснения особенностей его генетической основы достаточно плодотворными оказываются простые межлинейные сравнения и анализ поведения гибридов.
Ниже мы приведем короткую сводку результатов генетических исследований ориентировочно-исследовательского поведения, агрессивности и способности к обучению, проведенных традиционными методами, применявшимися в генетике поведения в течение всего периода ее формирования — в 60—80-е годы.
Роль отдельных генов в формировании некоторых признаков поведения будет рассмотрена на примере гена альбинизма, генов, мутации по которым вызывают неврологические нарушения, а также генов, экспрессирующихся у трансгенных животных.
Исследовательское поведение, вызываемое новой обстановкой и новыми предметами, представлено у грызунов поведенческими актами и позами, которые способствуют сбору информации о незнакомых элементах ситуации.
Как упоминал ось выше, существование генетического компонента изменчивости исследовательского поведения проиллюстрировал X. Ван-Абилен (van Abeelen, 1974). У мышей двух инбредных линий C57BL/6J и DBA/2J число вертикальных стоек в установке "открытое поле" достоверно различалось. Как показал анализ гибридов первого и второго поколения, этот типично количественный признак имеет полигенный тип наследования. На основе популяции межлинейных гибридов была начата селекция мышей на высокие и низкие значения признака — числа стоек в ситуации "открытого поля". В результате этого эксперимента еще в начале 70-х годов были получены линии, достоверно различающиеся по уровню исследовательской активности. Они представляли собой удобную физиолого-генетическую модель для исследования особенностей мор фологической и биохимической организации мозга, обеспечивающих различия в этом виде поведения мышей.
Одним из направлений таких исследований был сравнительный анализ состояния холинергической, ГАМКергической и опиатной систем гиппокампа у инбредных линий C57BL и DBA, а также у двух селектированных линий. Различающиеся по поведению линии давали и противоположно направленные реакции на введение ряда фармакологических веществ, воздействующих на упомянутые выше мозговые медиаторные системы. Внутримозговое (в гиппокамп) введение скополамина — антагониста холинергической системы — снижало показатели исследовательской активности у мышей C57BL и усиливало их у DBA, т.е. инвертировало различия, наблюдаемые в норме. Сходные результаты были получены и на селектированных линиях.
Очевидно, что адаптивная реакция на новизну — исследование обстановки — предполагает сбалансированное изменение активности ряда нейронных систем мозга, в частности гиппокампа. Ван Абилен предположил, что у линии C57BL такая согласованность более устойчива, чем у DBA. Роль особенностей структуры гиппокампа в формировании поведения рассмотрена также в разделе 8.6.7.2.
Поведение животных в установке "открытое поле" было предметом многих генетических исследований. Поскольку помещение крысы и/или мыши на открытую освещенную площадку вызывает не только стремление исследовать новую обстановку, но и страх, то традиционно показатели поведения в этой ситуации принято трактовать как динамический баланс этих двух тенденций. Поэтому высокий уровень локомоции (как правило, при низком уровне эмоциональной реактивности) принято трактовать как показатель высокой исследовательской тенденции (хотя во многих работах в качестве собственно исследовательских движений рассматривались лишь стойки), тогда как обратное соотношение этих показателей обычно служит признаком высокого уровня пугливости.
В период интенсивного накопления фактов генетикой поведения были проведены селекционные эксперименты по выведению линий, контрастных по названным выше признакам. Одна из этих работ — селекция мышей на разные уровни двигательной активности в "открытом поле". Другой известный эксперимент, который был одним из первых в данной области, — это выведение английским ученым П. Бродхерстом (1975) двух линий крыс (Maudsley Reactive и Maudsley Nonreactive strains), контрастных по уровню эмоциональной реактивности (оценивалась по числу актов дефекации) в "открытом поле". В дальнейшем подобные селекционные эксперименты на крысах и мышах проводились неоднократно, в результате чего был установлен ряд существенных фактов. Так, разные уровни активности и разная "эмоциональность" оказываются как бы в антагонизме друг с другом. Иными словами, отбор на контрастные значения одного признака, ведет к появлению противоположных по уровню, но также достаточно выраженных различий по другому признаку. Высоко активные животные оказываются "малоэмоциональными" и наоборот. В истории исследования поведения грызунов разных видов и групп, различающихся по полу, возрасту и функциональному состоянию, имеется много примеров, когда этот антагонизм отсутствует, и отрицательная корреляция между уровнями двигательной активности и эмоциональной реактивности не является строго обязательным явлением.
Следует отметить, что в этих селекционных экспериментах также были выявлены и плейотропные эффекты, и коррелированные ответы на отбор. Одной из особенностей линий Maudsley, важной для физиологической интерпретации данных, является более развитая иннервация кишечника (за счет энтеральной нервной системы) у крыс, которые в ситуации "открытого поля" совершали большее количество актов дефекации. Это может означать, что интерпретировать высокий уровень дефекации у Maudsley Reactive исключительно как показатель высокой эмоциональной реактивности было бы неверно (Blizzard et al., 1982).
Для оценки ориентировочно-исследовательского поведения лабораторных животных применяются разные экспериментальные установки и методы, что может быть причиной наблюдаемых различий в поведении заведомо стандартных линий мышей и крыс. Нередко результаты работ разных авторов даже противоречат друг другу. Как правило, это не служит препятствием в проведении дальнейших исследований, а наоборот, позволяет выяснить возможные причины различий.
При исследовании форм поведения млекопитающих и анализе дифференциального участия в них некоторых отделов мозга все большее значение начинают приобретать так называемые фармако-генетические исследования. Речь идет об оценке реакций на известные (или новые) фармакологические препараты или химические агенты у животных разных генотипов. Сильно упрощая картину, эти работы можно подразделить на несколько направлений: 1) оценку генетической изменчивости в продукции того или иного метаболита, участвующего в осуществлении какой-то реакции; 2) определение изменчивости числа и строения молекул-рецепторов; 3) исследования генетических особенностей эффекторной реакции и др. Одной из "опасностей" фармако-генетических исследований является тенденция перехода на все более простые объекты — срезы мозга, культуру клеток, фракции синаптосом и т.д. С одной стороны, создаются условия для выявления молекулярных механизмов поведенческих реакций, но с другой — затрудняется общая оценка роли таких факторов в формировании поведения. Ввиду этого особо важное значение имеют исследования, проводимые на животных с разными генотипами, а также отслеживание формирования того или иного признака в ходе онтогенеза.
Необходимо отметить и еще одно направление исследований в генетике поведения. Анализ генетической изменчивости некоторых поведенческих реакций, например исследовательской активности мышей (см.: 8.6.6.1), позволяет делать общие выводы об эволюции этой формы поведения.
Всякому, кто изучает поведение животных, очевидно, что термин "агрессивность" относится в целом к поведению животных при конфликтах (см.: 5.3.2). Многие исследователи используют также альтернативный термин — "агонистическое поведение", под которым понимают такие поведенческие акты, которые, имея общую задачу адаптации к обстановке, предполагают, тем не менее, физический конфликт между особями одного вида. Существуют разные формы таких конфликтов: это агрессия хищника, межсамцовые конфликты, материнская агрессия, агрессивное поведение в ситуации, вызывающей страх (Попова^ 1988). Такое поведение называется также активно-оборонительной реакцией, ее генетические основы исследовались Л.В. Крушинским (1991) на собаках еще в конце 30-х годов.
Агрессия хищника. Агрессия хищника — это одновременно и специфическое явление, связанное с пищевым поведением, и проявление агонистического поведения, имеющего общие черты с агрессией других типов. Агрессию хищника изучают на разных моделях — на крысах, убивающих мышей и сверчков, на мышах, "охотящихся" на сверчков и саранчу, на пушных зверях — норках и лисицах, у которых анализируется реакция на традиционную для них добычу.
Генетические исследования обнаружили не только межлинейные различия, но и доминирование у мышей высокого уровня агрессии хищника. Данные генетического анализа предположительно свидетельствуют о наличии у мышей двух главных генов, которые контролируют признак "агрессия хищника".
Агрессивность, адресованная человеку, — весьма специфический тип поведенческих реакций. Она была объектом специальных исследований большого коллектива ученых под руководством академика Д.К. Беляева (см. выше).
В длительных селекционных экспериментах, проведенных в Институте цитологии и генетики СО АН СССР в Новосибирске, были получены линии серебристо-черных лисиц и серых крыс, различающиеся по уровню агрессивности, возникающей при контакте с человеком. В последние годы выведены такие же линии норок. Heагрессивная линия лисиц, полученная в результате отбора, обладает необычным для этого вида типом поведения: они активно ищут контакта с человеком и не испытывают ни малейшего страха перед ним. Линии крыс, селектированные на основе выборки отловленных в природе крыс-пасюков, также резко отличаются по агрессивности, адресованной человеку.
Специальные эксперименты показали, что крысы, агрессивные в отношении человека, не показывают столь же выраженной агрессии хищника. К 20-му поколению селекции крыс на высокую и низкую агрессивность по отношению к человеку у них не произошло изменений в уровнях агрессии хищника и межсамцовой агрессии. Это означает, что "агрессия хищника" и "агрессия к человеку" имеют разные физиолого-генетические механизмы.
Нейрохимические исследования показали, что отбор на разные уровни агрессивности, адресованной человеку, позволяет получить линии с измененным гормональным статусом, специфика которого проявляется как в состоянии покоя, так и в состоянии испуга и стресса.
Анализ особенностей поведения животных в условиях их "принудительного" контакта с человеком привел Д.К. Беляева к выводу, что агрессия серебристо-черных лисиц, направленная на человека, сопровождается формированием у них стрессорной реакции, а отбор на тип поведения "ручные животные" (одомашнивание) фактически означает формирование стрессоустойчивого генотипа (см. также: 8.6.5.3).
Межсамцовые конфликты. Как известно, у самцов многих видов существуют две формы поведения при социальном конфликте — нападение и защита (см. также: разделы 4.3 и 5.3.2). Они различаются как по характеру движений (наборам ФКД) животных, так и по "мишеням" (особенностям нанесения укусов).
Для грызунов характерны преследование противника, вертикальные и боковые позы угрозы, укусы с отталкиванием. Укусы нацелены главным образом на спину, бока и основание хвоста соперника. При защите животное ведет себя по другому: для него характерны главным образом реакция бегства, позы "защиты" и "подчинения", а также нанесение укусов "в прыжке".
Генетические исследования реакции нападения практически всегда осуществляются на основе теста "попарного ссаживания" двух самцов. Существуют три типа таких "диадных" тестов.
Во-первых, можно сажать вместе животных одного и того же генотипа, например одной линии или гибридов первого поколения. Во-вторых, можно исследовать конфликт между животными разных генотипов. Третий способ — это метод "стандартного тестера", когда конфликты индуцируются между животными анализируемого генотипа и определенного стандартного генотипа. Этот самец, стандартный тестер, не должен сам инициировать конфликт (нападать), но при этом от него требуется способность спровоцировать реакцию нападения у партнера, не "давая сдачи". Часто в качестве стандартного тестера используют кастрированных гибридных животных, на тело которых наносят капли мочи самцов нужного исследователю генотипа. Иногда используют самцов с выключенным обонянием (либо путем удаления обонятельных луковиц, либо путем орошения обонятельного эпителия сульфатом цинка), что снижает их агрессивность.
Данные, получаемые при тестировании разными методами агрессивной реакции нападения, нередко различаются. Так, например, у самцов линии BALB/c число драк было выше, чем у C57BL, когда их тестировали с самцами той же ("своей") линии; при использовании метода стандартного тестера соотношение интенсивности реакций нападения было обратным. Это означает, что данная простейшая форма социального поведения определяется не только генами данной особи, но и спецификой раздражения (зрительного, обонятельного и др.), идущего от оппонента, т.е. также в значительной мере определяется его генотипом. Очевидно, что этот осложняющий исследование фактор следует непременно учитывать при генетическом анализе числа и интенсивности атак. Существенно важно, в какой обстановке — новой или же в привычной домашней клетке — происходит тестирование. В последнем случае этот тест носит название "резидент — чужак (интрудер)".
Химическая коммуникация, которой принадлежит важнейшая роль при формировании социальных контактов в сообществах грызунов, также находится под генотипическим контролем. Подробная и крайне поучительная сводка работ по данной проблеме была сделана С.Н. Новиковым в 80-х годах (Новиков, 1988), но не утратила актуальности и сейчас.
Следует отметить, что анализ генетических закономерностей межсамцовой агрессии проводят при "спонтанной" агрессивности, а также при агрессивности, резко усиленной содержанием животного в изоляции. Агонистическое поведение как признак несколько различается по своим свойствам при этих двух типах его индукции.
При исследовании генетики агрессивности оценивается, как правило, целый ряд параметров: латентный период первой атаки, число нападений за фиксированный интервал времени, длительность агонистического контакта, его интенсивность, а также доля агрессивных животных в группе.
Для всех перечисленных признаков характерна генетическая изменчивость, однако корреляции между числом самцов, проявляющих агрессию, и интенсивностью их агонистического поведения обнаружено не было. Другими словами, межлинейные различия в экспрессивности и пенетрантности разных показателей агонистического поведения носят сложный характер и не совпадают между собой. Специальные опыты показали, что эти признаки определяются разными генетическими системами. Такие признаки, как суммарное время, проведенное парой мышей в состоянии конфликта, и число дискретных атак наследуются аддитивно и не связаны с полом, тогда как в наследовании уровня агрессивности (доле агрессивных животных) доминирует низкий порог агрессивной реакции (иногда даже с эффектом сверхдоминирования). Вместе с тем, нельзя забывать, что в определении фенотипа агонистического поведения участвует много независимых переменных, следовательно, простых результатов при решении этой проблемы быть не может.
Сложная картина формирования агонистического поведения с позиций генетики означает, что исследователи должны уделять большое внимание поиску удобных, надежных и простых генетических моделей, которые позволяли бы свести к минимуму влияние внешних условий. Работа на таких моделях дает возможность использовать современные методы генетических исследований: методы рекомбинантных и рекомбинантных конгенных линий, картирование локусов количественных признаков, определение полиморфизма длины рестрикционных фрагментов ДНК и др.
Эксперименты, в которых проводили классический менделевский генетический анализ признаков, связанных с реакцией нападения, показали, что целый ряд ее характеристик генетически детерминирован, и их наследование можно описать как моно- или двухфакторное.
Межлинейные различия по особенностям самцовой агрессивности определяют в основном выбор доминанта в однополой, искусственно созданной группе самцов разных генотипов. Работы сотрудников Института цитологии и генетики СО АН СССР (Новосибирск) показали, что линия РТ, а также гибриды первого поколения линий РТ и СВА имеют устойчивую тенденцию поставлять доминантов, а целый ряд других инбредных линий располагаются в порядке убывания этой способности (Осадчук, Науменко, 1981).
Селекционные эксперименты с отбором мышей и крыс на разные уровни агрессивности предпринимались неоднократно. Широкую известность в этом плане получила работа К.И.Лагершпец и К.М.Лагершпец (К. Lagerspetz, К. Lagerspetz, 1974) по селекции крыс.
Ван Оортмерсен с сотр. (Benus et al., 1987; 1988) путем искусственного отбора создали две линии мышей, различающихся по агрессивности. Селекция проводилась из генетически гетерогенной популяции диких мышей. В качестве критерия для отбора был взят латентный период первого нападения (при тестировании со стандартным тестером — самцом линии MAS-Gro). Латентные периоды менее 200 сек считались короткими, более 200 сек — длинными. Уже через 8 поколений величины латентных периодов первой атаки у двух селектированных линий практически не перекрывались. Отбор в этой работе проводили по поведению самцов, а тестирование самок на агрессивное поведение не выявило межлинейных различий. Интересно отметить, что селекция самок диких мышей на высокую и низкую агрессивность не привела к различиям в агрессивности самцов полученных линий. Это еще раз подтверждает, что, несмотря на внешнее сходство двигательных актов агонистического поведения разного вида, контролирующие их генетические системы оказываются, по крайней мере частично, разными, и могут зависеть от пола.
Видоспецифические движения и позы при агрессивных реакциях, будь это нападение матери, защищающей гнездо, или межсамцовый конфликт, достаточно сходны между собой. Это свидетельствует об определенной общности физиологических механизмов обеих реакций. Однако подобное сходство внешней формы не должно заслонять от нас того обстоятельства, что генетические закономерности проявления разных агрессивных реакций (например, агрессии хищника, межсамцовой агрессии и др.) могут сильно различаться, как об этом уже говорилось выше. Более того, в зависимости от критерия отбора на агрессивность даже линии, полученные в результате отбора, также могут иметь разную "структуру" агрессивного или неагрессивного фенотипа. Линии Turku agressive и Turku nonagressive (К. Lagerspetz, К. Lagerspetz, 1974), как и линии Ван-Оортмерсена, различавшиеся по межсамцовой агрессивности, имели и разные уровни материнской агрессии. Критерий отбора в этой работе был иным: при тестировании самцов для отбора их не содержали в изоляции. Такие различия в особенностях фенотипического проявления агрессивности у разных линий могут быть использованы для изучения физиологических механизмов агрессивного поведения.
На поведение животных агрессивных и неагрессивных линий могут оказывать большое влияние факторы внешней среды, такие как опыт предыдущих конфликтов, воздействия на животных в ходе онтогенеза, изменение гормонального фона особи.
Уровни секреции гормонов, в частности тестостерона, также находятся под контролем генотипа. Генетическая система, "ответственная" за этот признак, связана с Y-хромосомой. В какой степени генетическая регуляция уровня тестостерона, а следовательно, и уровня половой активности самцов, связана с системой генетического контроля межсамцовой агрессивности?
Интерес к проблеме вызвал появление серии оригинальных исследовании роли генов, локализованных в Y-хромосоме, при формировании агрессивного фенотипа самца мыши (Maxson, 1992). Как известно, в Y-хромосоме выделяют два участка. Один из них называется псевдоаутосомным. Он может рекомбинировать в мейозе с X-хромосомой, располагается в теломерном участке длинного плеча Y-хромосомы, и передается как дочерям, так и сыновьям данного самца. Другой, истинно моносомный (т.е. специфичный для половой хромосомы) участок, передается строго от отца к сыну и называется непсевдоаутосомным. Он состоит из короткого плеча, центромерного района и участка длинного плеча Y-хромосомы вплоть до псевдоаутосомного участка. Непсевдоаутосомный участок содержит гены, ответственные, например, за детерминирование развития гонад в семенники, ген, кодирующий H-Y антиген, и др. Поскольку этот участок может передаваться только от отца к сыну, разработаны системы скрещиваний, позволяющие определять, влияют ли расположенные в нем гены на реакцию нападения. Можно, например, сравнить реакцию нападения у самцов—реципрокных габридов первого поколения от скрещивания линий грызунов, контрастных по данному признаку. Если поведение таких самцов-детей и их отца окажется сходным, то это можно истолковать как свидетельство участия одного или более генов непсевдоаутосомного участка в формировании признака (см. также: Carlier et al., 1990).
Тщательные исследования поведения мышей, у которых Y-хромосома имела происхождение либо от линии DBA/1, либо от C57BL/10, показали, что изменчивость агрессивного поведения самцов связана с группой генов псевдоаутосомного участка Y-хромосомы, т.е. той части генома, которая рекомбинирует с X-хромосомой самки. Этот участок контролирует длительность латентного периода первой атаки у линий Ван Оортмерсена. Возможно, что подобная локализация по крайней мере ряда генов, связанных с агрессивностью, объясняет некоторую взаимозависимость агрессивных реакций, проявляющихся в разных ситуациях.
Межсамцовая агрессивность при жизни мышей в естественных условиях есть фактор, обеспечивающий установление иерархии внутри небольших сообществ, из которых собственно и состоит популяция домовой мыши (см.: 5.5.1). Установившаяся внутригрупповая иерархия обеспечивает минимальный уровень агрессии в "повседневной жизни" групп, а также служит фактором, определяющим расселение молодых половозрелых самцов на новые территории.
Способность к обучению — это наиболее трудная, но и наиболее увлекательная область генетики поведения. Первая последовательная попытка селекции крыс на способность к обучению (опыт Трайона) показала практически все трудности, подстерегающие исследователя на этом пути.
Селекция крыс на успешность (количество ошибочных заходов в тупики) обучения поиску пищи в лабиринте (рис. 8.22) была одной из первых работ по генетике поведения (см.: 8.6.2). Было показано, что крысы "умной" линии ТМВ обучались значительно лучше в исходном 17-тупиковом лабиринте, а также в более простом, 14-тупиковом. В то же время при обучении в 16- и 6-тупиковых лабиринтах межлинейных различий обнаружено не было.
Рис. 8.22. Селекция крыс на высокий и низкий уровень обучаемости (число ошибок) в многотупиковом лабиринте (опыт Трайона). На а: 1 — родительское поколение; 2 — 4-е поколение селекции; 3 — 8-е поколение селекции; на б: 1 — "умные" крысы; 2 — "глупые" крысы
Обучение реакции избегания воды было более успешным у "глупой" линии TMD. В дальнейшем было показано, что ТМВ лучше обучались реакции активного избегания в челночной камере, но в тесте на обучение избеганию тока, когда надо было выпрыгивать из (а не переходить из одной ее половины в другую) показатели TMD были выше. Это может означать, что в эксперименте Трайона отбор животных производился (неосознанно, разумеется) не только на способность к обучению, но и на какие-то особенности поведения, связанные с лабиринтом данной конфигурации. У крыс линии ТМВ выше пищевая мотивация, но слабее оборонительная, они меньше отвлекаются при выполнении навыка, тогда как крысы линии TMD пугаются даже при манипуляциях дверцами лабиринта. Исследователи приходят к выводу, что ТМВ лучше решают тесты, связанные с пространственными стимулами, тогда как TMD успешнее обучаются при зрительных раздражителях.
Другие формы поведения трайоновских линий в значительной степени различаются между собой, причем физиологическая интерпретация различий иногда достаточно проста, иногда затруднительна или просто невозможна. У линии ТМВ был ниже порог провокации ЭЭГ-знаков судорожной активности при электростимуляции миндалины, слабее реакция на новые предметы, ниже уровень локомоторной активности в колесах. Можно полагать, что отбор на разную степень успеха обучения в лабиринте способствовал формированию генотипов, при которых особенности процесса восприятия, параметры пространственной памяти, мотивация и другие фенотипические признаки оказались в одном случай оптимальны, а в другом — субоптимальны для выполнения данной реакции.
Результаты сравнения поведения трайоновских линий достаточно поучительны в том отношении, что иллюстрируют сложную картину, которая может получиться в результате отбора на способность к обучению, если реально отбору подвергается признак, косвенно связанный с изучаемым. Несомненно, что при создании путем отбора трайоновских линий произошло видоизменение их поведенческих фенотипов. В то же время достоверного прямого влияния отбора на собственно способность к обучению не обнаруживается. Примером сложности полученной картины является изменение способности к обучению крыс этих линий при выращивании их в условиях с разной степенью "обогащенное'' среды (рис. 8.23; см. также: 7.3).
Рис. 8.23. Исчезновение различий и изменение уровней выполнения навыка у крыс линий Трайона в условиях нормальной, "обедненной" и "обогащенной" среды. По оси ординат — число ошибок при обучении (Мак-Фарленд, 1988)
Реакция активного избегания удара электрического тока в челночной камере — это четкий, легко поддающийся количественному учету тест на обучаемость у лабораторных грызунов. Поскольку в качестве условного раздражителя можно выбрать звук, то при использовании этого теста можно включать в сравнение и животных-альбиносов, не опасаясь, что низкая острота их зрения повлияет на успешность выполнения навыка. Челночные камеры, в которых животные обучаются попеременно переходить из одного отсека в другой при предъявлении условного сигнала, легко автоматизировать, что способствует получению данных на большом числе животных в унифицированных условиях. Все это сделало данную методику очень популярной среди исследователей, а число работ, выполненных с ее помощью, составляет многие сотни. Особенно велико число фармакологических и фармакогенетических исследований.
В начале 60-х годов на основе популяции крыс Вистар итальянский исследователь Дж. Биньями начал селекцию на высокую и низкую способность крыс к обучению реакции активного избегания в челночной камере. Уже через несколько лет эти линии были сформированы. С начала 70-х годов селекция и разведение этих крыс проводились независимо в Швейцарии, Великобритании и Канаде.
Межлинейные различия по селектируемому признаку высокодостоверны. Эти линии получили название римских (Roman High Avoidance, RHA, Roman Low Avoidance, RLA) и интенсивно исследуются во многих лабораториях мира.
Сравнение результатов огромного числа работ, полученных на этих линиях с использованием практически всех существующих методик оценки поведения, привело исследователей к заключению, что межлинейные различия у RHA и RLA связаны с эмоциональностью и разным типом реакции на стрессоры (Жуков, 1997; см. также: 8.6.5.3). В то же время существуют достаточно убедительные данные, что эти линии показывают различия и в ассоциативных способностях, т.е. в способности к обучению как таковой. Большое внимание нейроэндокринологическим механизмам различных стратегий поведения крыс в условиях избегаемого и неизбегаемого удара электрического тока было уделено в работах на крысах двух селектированных линий — КНА и KLA (Koltoushi High Avoidance, Koltoushi Low Avoidance), иначе называемых линиями ВЛ и НЛ. Выведение этих линий было начато в конце 50-х годов в Институте физиологии им. Павлова АН СССР В.К.Федоровым, а после его смерти продолжено Н.Г.Лопатиной с соавт. (см.: Лопатина, Пономаренко, 1987). Как и другие линии крыс, селектированные на высокую и низкую способность к избеганию в челночной камере, данные линии отличаются друг от друга и по иным признакам, связанным как с поведением, так и с более "простыми" физиологическими реакциями. Подробный анализ экспериментальных данных, полученных на крысах этих линий, проведен в монографии Д.А.Жукова (1997).
Способность крыс селектированных линий к обучению реакции избегания была сопоставлена с таким морфологическим показателем, как размер интра- и инфрапирамидных проекций мшистых волокон — аксонных окончаний нейронов зубчатой фасции в поле СА3 гиппокампа. Эта проблема рассмотрена более подробно в разделе 8.6.7.2.
Позднее, уже в 70-е годы были созданы путем селекции еще две линии (Syracuse High Avoidance, SHA, Syracuse Low Avoidance, SLA), различающиеся по обучению реакции активного избегания, но не обнаруживающие, в отличие от римских линий, различий в уровне двигательной активности. Исходной популяцией для этой селекционной работы служили крысы Лонг-Иванс. В настоящее время крысы хорошо обучающейся линии SHA дают примерно 40 реакций избегания в 60 предъявлениях теста, тогда как SLA — ни одного. У SLA сильно подавлены межсигнальные реакции, однако скорость выполнения реакций избавления (т.е. реагирования на включение электрического тока-наказания) у них не отличалась от линии SHA. В тесте "открытого поля" SLA, как и RLA, более эмоциональны, однако различий в уровнях локомоции у них, как сказано выше, не выявлено.
Генетические модели способности к обучению, какими являются линии крыс RHA и RLA, а также SHA и SLA, используются также в исследованиях роли наиболее изученных медиаторных систем в формировании межлинейных различий реакции на стресс, в обучении, в организации болевой чувствительности. Эти исследования представляют большой интерес, но в настоящем пособии не могут быть затронуты сколько-нибудь подробно (см.: Gray, 1987),
Другим аспектом проблемы, который делается очевидным при анализе межлинейных различий в обучении реакции избегания в челночной камере, является биологическая адекватность самого теста. В самом деле, анализ "логики" этого теста показывает, что данная реакция далека от того, чтобы имитировать какую-либо из естественных поведенческих реакций крысы или мыши. Животное, находящееся в камере, получает болевое раздражение (удар тока), которому предшествует некий сигнал (свет и/или звук). Бурная двигательная реакция в ответ на боль "переносит" крысу или мышь в другую половину камеры, где ток (и наказание) отсутствует и где она получает отдых. Однако через короткое время в этом бывшем безопасном отсеке животное получает новый удар тока, а метания по клетке переносят его в отсек, где ранее также предъявлялось наказание. В этих условиях у животных (тем не менее) формируется условная реакция выполнения перехода из одного отсека в другой в ответ на предъявление условного сигнала (света или звука).
Реакция перехода выполняется в двух половинах камеры (в двух разных контекстах), причем пребывание в каждом из них регулярно наказывается (если реакция не выполняется). Это означает, что в данном тесте животное должно искать убежища там, где оно только что получило наказание.
Возможно, эта логическая противоречивость теста и есть причина того, что множество генетических линий и групп лабораторных грызунов демонстрирует весьма невысокие показатели обучаемости, хотя у некоторых животных они, напротив, высокие. В чем же заключаются различия между такими животными? Ответ на этот вопрос исследователи нашли с помощью генетико-этологического анализа проблемы (Вилкок, 1975). Генетический эксперимент, который позволил найти путь его разрешения, изложен в разделе, посвященном описанию методов диаллельного скрещивания (см.: 8.6.6.2).
Этологический аспект проблемы заключается в следующем. Боль как фактор, провоцирующий у животного реакцию страха, вызывает у млекопитающих, а точнее у грызунов, одну из двух ответных реакций: либо замирание (затаивание), либо бегство.
Существует так называемая двухкомпонентная теория физиологических механизмов реакции избегания, в соответствии с которой в начальный период обучения выявляется тенденция либо "затаиваться" от опасности, которую представляет собой боль, предваряемая условным сигналом, либо убегать, но обе реакции сначала осуществляются вне связи с условным сигналом. Преобладание у животных реакции "затаивания" ухудшает их шанс усвоить данный навык. В целом, по мере увеличения числа сочетаний условного и безусловного сигналов в поведении начинает превалировать тенденция к бегству. Это не объясняет, почему смена реакций обнаруживает подобную динамику, однако для анализа поведения в челночной камере с позиций генетической изменчивости объяснение представляется продуктивным.
При отборе животных в процессе выведения линий, быстро и медленно обучающихся условнорефлекторной реакции избегания, исследователь руководствуется условными критериями обученности, точнее, тем, как быстро животные, которые будут использованы в качестве производителей, их достигают. Можно предполагать, что крысы, быстрее достигшие данного критерия, действительно обладают более высокими ассоциативными способностями. Однако естественно предположить и другое: исследователи отбирают не только "умных" и "глупых" животных, но и таких, у кого оказываются более низкими пороги реакции бегства в ответ на боль (более высокий уровень реакций избегания), а также тех, у кого ниже пороги реакции замирания как ответа на боль (уровень реакций избегания ниже). Данные опыта по диаллельному скрещиванию (см.: 8.6.6.2) в целом подтверждают правомочность такого предположения.
Как уже упоминалось, в настоящее время разработаны экспериментальные приемы, с помощью которых в геном животного можно ввести последовательность оснований, кодирующую определенный белок. Этот ген может быть новым для вида или видоизмененным геном вида-хозяина. В последнем случае, как правило, речь идет о "выключении" какого-либо гена из процесса развития. Организм такого трансгенного животного (это как правило, мышь или дрозофила) развивается в новых условиях, когда данный ген не может экспрессироваться нормально.
Экспериментальные схемы обучения, которые используются для тестирования запоминания у лабораторных мышей и крыс, позволяют с большой надежностью разделить влияние какого-либо фактора на краткосрочную и долгосрочную память, а также на процесс собственно усвоения навыка. В основе такого навыка обычно лежит простая двигательная реакция или, наоборот, ее торможение (невыполнение). Усвоение подобного навыка происходит при единственном сочетании условного и безусловного раздражителей, что также важно для четкости оценки эффекта мутации.
Многочисленные данные об участии системы вторичных посредников в формировании следа памяти позволяют считать доказанным, что долговременная память связана с изменениями в структуре синаптических белков, причем эти изменения осуществляются в результате целого каскада событий, одним из которых является активация гена, кодирующего белок CREB. Для выяснения его роли в формировании памяти были получены мыши, у которых отсутствовал ген, кодирующий белок CREB (Bourchaladze et al., 1994; см.: 8.4.3.6). В интервалах "работы" краткосрочной памяти (30 и 60 мин после сеанса обучения) запоминание навыка было достоверным, тогда как при тестировании в сроки, когда должна "работать" долгосрочная память (через 2 ч), воспроизведение навыка было сильно нарушено.
Было продемонстрировано также, что феномен долговременной потенциации, который многие рассматривают как гомолог условного рефлекса, формирующийся на уровне нейронных ансамблей гиппокампа, у мышей с отсутствием гена, кодирующего белок CREB, развивался аномально в тех же временных пределах. Через 2 ч после воздействия, вызывающего долговременную потенциацию в срезах мозга (гиппокампе) таких животных, все ее проявления уже отсутствуют.
Мыши с искусственной мутацией гена калмодулин-зависимой протеинкиназы II (Bach et ah, 1995) нормально обучались навыку отыскания безопасного убежища при наличии сигнальных раздражителей, но не могли усвоить этот навык, когда для этого требовалось формирование пространственного представления. При этом у них наблюдалась еще одна особенность: при раздражении с частотой 5—10 в секунду (т.е. с частотой тета-ритма, как правило, присутствующего в электрограмме гиппокампа при исследовательском поведении) долговременной постсинаптической потенциации не было, в то время как при высокочастотном раздражении гиппокампа она развивалась нормально.
Таким образом, совокупность данных, полученных на животных разного уровня организации, позволяет в настоящее время считать, что экспрессия транскрипционного фактора CREB, который активирует гены, прямо связанные с формированием памяти, и ряд других генетических элементов, как правило, связанных с функцией системы вторичных посредников, являются важным этапом записи следа памяти в мозге.
В разделе 8.4.3.6 приводились примеры влияния мутаций отдельных генов на разные формы поведения дрозофилы. Эффекты отдельных генов на поведение млекопитающих (преимущественно мыши) бывают не менее иллюстративными. Оценка влияния одиночных генов на поведение требует глубоких знаний по физиологии поведения, поскольку в противном случае интерпретация результатов может быть ошибочной. Данные по роли гена "albino" в поведении лабораторных грызунов в этом отношении весьма поучительны.
Альбинизм (депигментация) — это мутация локуса с, локализованного у мышей на 7-й хромосоме. Мутация рецессивна и вызывает целый ряд других аномалий, в частности в зрительной системе. Показано, например, что у альбиносов многих видов происходит практически полный перекрест волокон в хиазме, т.е. у них отсутствуют ипсилатеральные зрительные проекции.
Межлинейные сравнения. В поведении животных-альбиносов также обнаруживается ряд особенностей. Как показали сравнительные исследования частоты самораздражения мозга у мышей линий BALB/c и DBA/2, средняя частота нажатий на рычаг для раздражения зон "удовольствия" в латеральном гипоталамусе у мышей BALB/c была почти в два раза выше, чем у DBA/2. Чтобы выяснить вопрос, в какой степени эти различия связаны с отсутствием пигментации у BALB/c, был проведен классический генетический анализ. В специальных скрещиваниях были получены гибриды первого и второго поколения, а также беккроссы — потомки от скрещивания гибридов с родительскими формами. Среди поколения бек-кроссов, у которых произошло расщепление по окраске шерсти, среди мышей с высокой и низкой частотой самораздражения пигментированные животные и альбиносы были представлены с равной частотой. Таким образом, связь между частотой самораздражения и альбинизмом оказалась случайной, а не функциональной, и ассоциация исчезла при более подробном анализе признаков. Этот пример является одной из многочисленных иллюстраций общего правила необходимости анализа обнаруженных межлинейных различий в последующих экспериментах по скрещиванию и расщеплению в гибридных поколениях.
У особей-альбиносов всех видов (включая и человека) отсутствует меланин, причем не только в коже и волосах, но и в радужной оболочке глаз. Отсутствие пигментации глаз (и соответственно измененная зрительная функция) у мышей и крыс-альбиносов требовало специального внимания при проведении сравнительных исследований по генетике поведения грызунов, в частности по методу "открытого поля".
Мышей линии C57BL/6J (имеющих черную шерсть, высокий уровень активности и низкий уровень дефекации в установке "открытое поле") скрещивали с белыми мышами линии BALB/c (с низким уровнем активности и высокими показателями дефекации). От гибридов первого поколения получили гибридов второго и третьего поколений, после чего сопоставили показатели активности и эмоциональности с наличием пигментации. Оказалось, что, как и у исходных линий, у альбиносов уровень активности был ниже, а уровень дефекации выше, чем у пигментированных животных. Получив третье поколение гибридов, американский исследователь Дж.Де Фриз, приступивший к этой работе в конце 60-х годов, начал селекцию на высокий, средний и низкий уровни активности в установке "открытое поле", не обращая внимания на цвет шерсти животных. Уже к 8-му поколению в линии, селектированной на высокий уровень активности, практически не было альбиносов, тогда как в линии с низкой активностью их было 100%. В другой паре линий, также селектированных из той же популяции на разные значения признака (уровень активности), низкоактивная линия на 60% состояла из непигментированных мышей. Эти полученные данные нельзя было объяснить сцеплением (близким топографическим расположением на хромосоме) двух локусов — локуса альбинизма "с" и какого-то другого, влиявшего на уровень активности, — поскольку низкая активность альбиносов сохранялась и после многих поколений неинбредного разведения, когда такая ассоциация могла многократно нарушиться вследствие кроссинговера. Кроме того, в проводимых параллельно двух селекционных экспериментах (т.е. при параллельной селекции еще трех линий на низкие, средние и высокие значения признака) результаты были сходными. Важность оценки таких коррелированных ответов на отбор достаточно велика, причем для селекционных экспериментов с любыми объектами.
Другая возможность объяснения заключалась в постулировании тривиального плейотропного эффекта гена альбинизма, который заключался в том, что подавление двигательной активности альбиносов было связано с их неспособностью адаптировать зрение к яркому освещению площадки и нормально перемещаться в этих условиях. И действительно, тестирование мышей этих селектированных линий при красном свете практически нивелировало различия в уровне активности.
Метод коизогенных линий. Другим достаточно точным методом, который позволил оценить влияние гена альбинизма на поведение и был минимально связан с возможными эффектами сцепления, оказался так называемый метод коизогенных линий, т.е. инбредных линий, отличающихся друг от друга только по мутации в данном локусе. Так, например, при разведении мышей линии C57BL/6J в ней была обнаружена мутация альбинизма, и оказалось возможным сравнить поведение мышей двух линий, которые были генетически идентичны друг другу за исключением гена "с". Даже в условиях тестов, где не было подавляющего влияния яркого света, мыши-альбиносы были менее активны в открытом поле и медленнее избегали воды. При тестировании в течение 90 мин в первые минуты теста активность альбиносов была достоверно более низкой, однако к концу тестирования различия исчезли. Уровень двигательной активности у этих групп мышей при тестировании в течение 24 ч в специальных колесах был одинаковым.
Метод конгенных линий. Использование в генетике поведения метода конгенных линий также связано с локусом альбинизма "с". Метод заключается в том, чтобы путем последовательных скрещиваний ввести в генотип определенной линии участок хромосомы, несущий интересующий экспериментатора локус. После этого возможно сравнение поведения животных, имеющих "новый", мутантный локус, с носителями исходного генотипа. Чтобы его сохранить, в каждом поколении новой линии проводят тестирование на наличие у мышей специфического аллеля тканевой совместимости. Это достигается использованием методики трансплантации участков кожи от мыши линии-носителя изучаемого хромосомного локуса. Локусы, обеспечивающие тканевую совместимость, хорошо описаны, картированы и достаточно равномерно распределены по геному, поэтому очень многие мутантные аллели, которые переносятся путем скрещиваний в другой генотип, можно маркировать именно генами гистосовместимости, относящимися к разным группам этой системы.
Таким же способом можно маркировать и ген "с", мутация по которому вызывает альбинизм. Эксперименты по скрещиванию мышей пигментированной линии С3Н с мышами-донорами локуса "с" проводились при постоянном тестировании потомства на наличие у них локуса гистосовместимости (по отторжению или приживлению кожного трансплантата от линии-донора). Таким способом было получено несколько линий, одна из которых оказалась депигментированной. Наблюдались также различия в уровне двигательной активности между исходной пигментированной линией С3Н и новой конгенной депигментированной линией. В начале тестирования альбиносы были достоверно менее активны, но к концу 90 мин теста различия исчезали.
Описанные методы применяются, разумеется, не только для выяснения роли альбинизма в поведении мышей. Они существенно важны для оценки роли генотипа в действии фармакологических агентов на поведение и на биохимические процессы. Использование метода рекомбинантных инбредных линий (см.: 8.6.6.3) делает результаты таких исследований еще более информативными (Demant et al., 1992).
Таким образом, помимо достаточно очевидного влияния депигментации на функцию зрения носители данной мутации по ряду признаков отличаются от пигментированных животных, но причина отличий не столь очевидна. Мутация альбинизма связана с изменениями (по некоторым данным, с делецией) структурного гена фермента тирозиназы. Это означает, что при развитии данного фенотипа могут происходить сложные компенсационные изменения, затрагивающие обмен, в частности таких важных веществ, как катехоламины. Подобные плейотропные эффекты могут быть причиной особенностей поведения животных-альбиносов. Эти вопросы еще предстоит исследовать экспериментально.
Еще одна категория мутационных изменений — хромосомные аберрации — также могут влиять на функции ЦНС. Из млекопитающих наиболее изученными в этом отношении оказались мыши.
Как известно, существует целый ряд хромосомных аномалий, возникновение которых в той или иной степени нарушает баланс физиологических процессов в организме. Наиболее известный и яркий пример трагического следствия подобного дисбаланса — так называемый синдром Дауна (см.: 8.6.5.6). В кариотипе мыши описаны практически все известные типы аномалий хромосом, достаточно подробно проанализировано их влияние на эмбриогенез (см.: Дыбан, Баранов, 1977), однако исследование поведения таких мутантов проводилось лишь в отдельных случаях.
В кариотипе мыши описано большое число хромосомных транслокаций*). Линии мышей, носителей мутаций этой категории, поддерживаются в лабораториях и питомниках, как например в Джексоновской лаборатории, наряду с другими инбредными мутантными линиями (см.: 8.1.1).
Влияние хромосомных аномалий на функцию ЦНС и поведение мышей одним из первых предложил исследовать отечественный эмбриолог проф. А.П.Дыбан (1976). На основании данных по специфическому влиянию такого рода аномалий на ход эмбриогенеза мыши он предположил, что характер влияния хромосомных перестроек на поведение может зависеть от типа хромосомной перестройки. Первой работой в этой области был анализ способности к обучению реакции активного избегания у мышей-трисомиков по маркерной хромосоме Т6, возникшей в результате транслокации — изменения строения хромосом 13 и 14 (Федоров и др., 1973). Мыши-трисомики, исследованные этими авторами, достоверно отставали в скорости приобретения навыка.
Далее, в совместной работе А.Л.Дыбана и B.C.Баранова с Л.В.Крушинским и его сотрудниками было проанализировано поведение мышей-носителей так называемых робертсоновских транслокаций**). Мыши-носители таких хромосомных перестроек заметно не отличаются ни по физическому развитию, ни по плодовитости. В лабораторных условиях их разводят так же, как животных обычных лабораторных линий.
*) Транслокация — разрыв одной или двух хромосом, вследствие чего участок одной хромосомы переносится на другую и/или образуется отдельная дополнительная хромосома. **) Робертсоновская транслокация — это слияние двух акроцентрических (те палочковидных) хромосом с образованием одной, мета- или субметацентрической. Цитогенетические исследования показывают, чго количество генетического мaтepиaла при этих транслокациях заметно не изменяется (хотя область околоцентромерного гетерохроматина, по некотррым данным, оказывается измененной)
Для анализа возможных особенностей поведения мышей с роберт-соновскими транслокациями был использован тест на способность к экстраполяции направления движения раздражителя, предложенный ранее Л.В. Крушинским (1986) для сравнения элементарной рассудочной деятельности животных разных видов, в частности крыс разного генотипа.
Суть методики заключалась в следующем: через небольшое отверстие в стенке камеры предварительно лишенное пищи животное начинало пить молоко. Кормушка с молоком, расположенная за этим отверстием, могла свободно перемещаться вправо или влево от мыши, причем ей был виден только начальный участок этой траектории. Иными словами, мышь получала информацию об исчезновении пищи и о направлении, в котором она исчезла. Задача заключалась в том, чтобы подойти к той стороне стенки, в направлении которой переместилась кормушка. Если доля правильных решений задачи (от всего числа предъявлений теста) достоверно отличалась от 50%-го (случайного) уровня, можно было делать вывод о неслучайном выборе "правильного" направления поиска. Следует сразу отметить, что подавляющее большинство тестированных мышей инбредных линий и аутбредных стоков давали в среднем именно 50%-й (т.е. случайный) уровень решения этого теста.
Способность к экстраполяции была проанализирована у значительного числа мышей, имевших в кариотипе робертсоновские транслокации разных хромосом. У животных Rb(6,15)lAld, Rb(9,14)6Bnr, Rb(16,17)7Bnr, Rb(5,19)1Wh (в скобках указаны номера хромосом, вовлеченных в слияние) доля правильных решений этого теста также не отличалась от 50%-го случайного уровня. В то же время при слиянии хромосом 8 и 17 (это были транслокации Rb(8,17)llem и Rb(8,17)6Sic, возникшие совершенно независимо и найденные в разных лабораториях) мыши были способны к экстраполяции (Крушинский и др., 1986). Они отличались и по способности к пространственной ориентации в лабиринте Морриса (см.: 8.6.7.2), но в данном тесте их показатели были достоверно хуже, чем у мышей с нормальным кариотипом (Leitingei et al., 1994). Специальные эксперименты показали, что такие отклонения поведения связаны с наличием в кариотипе именно данной робертсоновской транслокации, поскольку они встречались у мышей с разным генетическим фоном (Крушинский и др., 1986). Эта хромосомная мутация — слияние хромосом 8 и 17 — вызывает также изменения в содержании мозговых моноаминов, что, несомненно, свидетельствуют о ее влиянии на фунцкию ЦНС. По-видимому, феномен отражает так называемый эффект положения — изменение функции генетических элементов в зависимости от местоположения их в геноме. Нельзя исключить, что слияние хромосом изменяет их пространственное расположение в интерфазном ядре, а это, в свою очередь, может повлиять на процессы экспрессии некоторых генов.
Выше (см.: 8.5.4), при рассмотрении способов выявления первичного эффекта действия гена, мы касались проблемы нейрогенеза при неврологических мутациях. Вместе с тем, неврологические мутации интересны для генетики поведения и по ряду других причин. Генетические особенности поведения инбредных и селектированных линий мышей и крыс, а также мутантных линий и стоков с мутациями, затрагивающими отдельные звенья метаболизма, как правило, выражаются в виде слабых, хотя и достоверных отклонений. В то же время моногенные неврологические мутации, выявление которых стало возможным в условиях массового разведения мышей в питомниках, вызывают заметные отклонения в поведении. Эти мутации сопровождаются снижением жизнеспособности животных, у них сильно выражены отклонения в двигательной сфере, а морфология мозга или отдельных его структур в значительной степени изменена.
Все неврологические мутации — это моногенные нарушения. Наблюдения за мутантными животными позволяют составить "реестр" нарушений в двигательной и сенсорной сферах, а также нарушений поведения.
Поскольку одновременно проводится исследование первичных причин этих неврологических мутаций (см.; 8.5.4), сопоставление всего набора плейотропных эффектов первичного генетического дефекта с нарушением синтеза структурных или регуляторных белков на определенных стадиях развития поможет создать более полное представление о формировании, по крайней мере, самых простых функций мозга. В этом отношении неврологические мутации представляют собой потенциально мощный источник будущих знаний о мозге млекопитающих.
Внешне неврологические мутации проявляются в первую очередь в виде аномальной походки, трудностей в поддержании позы, нарушении способности к плаванию, исследовательскому поведению, в развитии судорожных состояний разной степени тяжести (Полетаева, Романова, 1990).
Первоначально эти дефекты связывали с нарушениями строения мозжечка и/или внутреннего уха, а также (у другой группы мутаций) с демиелинизацией мозга. Более подробные исследования позволили установить, что у таких мутантов нарушено строение продолговатого, среднего и переднего мозга, в том числе и гиппокампа.
Ниже приведен краткий и по этой причине неполный перечень аномалий поведения при неврологических мутациях мыши. Поскольку почти все неврологические мутации рецессивны, а гомозиготные по ним особи мало жизнеспособны (у них укорочена продолжительность жизни и резко снижена способность к размножению) мутантные линии поддерживают путем скрещивания между собой гетерозиготных мышей. При этом в потомстве обнаруживаются как гомо- и гетерозиготы, так и нормальные особи, которые обычно служат контролем при проведении исследований поведения мутантов.
Судорожные состояния. Судороги сопровождают мутации демиелинизации (например, quacking), а также weaver, Lurcher и tottering. Судороги в ответ на действие сильного звука (так называемая аудиогенная эпилепсия) обнаружены у мышей линии 101/HY, у которых мутация локализована в локусе, отвечающем за репарацию ДНК.
При мутации tottering (tg — ген локализован в 8-й хромосоме) наблюдается целый ряд аномальных фенотипических признаков. К их числу относятся повышенное содержание в мозге норадреналина и увеличение числа норадренергических терминалей нейронов голубого пятна (locus coeruleus — ядро продолговатого мозга). В другой мощной норадренергической системе мозга — в латеральных отделах покрышки (tegmentum) изменений не обнаружено. У гомозиготных особей tg/tg увеличено число адренергических аксонных терминалей нейронов голубого пятна на клетках новой коры, латерального коленчатого тела, гиппокампа, коры мозжечка, кохлеарного ядра. Ни размеры мозга, ни структура клеток, ни число норадренергических нейронов голубого пятна не отклоняются от нормы. В то же время у мутантов tg/tg периодически возникают эпилептиформные разряды в ЭЭГ, так называемые комплексы "пик-волна", а также судорожные припадки.
Более подробный анализ показал, что этот судорожный синдром, по-видимому, относится к категории типичных вторичных эффектов. Полагают, что повышенный уровень норадреналина в мозге этих мутантов вызывает компенсаторные изменения — снижение числа адренорецепторов на нейронах, что может повлечь за собой снижение чувствительности к норадреналину и далее — развитие судорог (см. также: 8.6.5.1).
Гнездостроительное и материнское поведение. Мышь-самка с мутацией staggerer (sg, 9-я хромосома, гибель почти 75% клеток Пуркинье и почти всех клеток-зерен) не поедает плаценту и не слизывает амниотические оболочки новорожденных мышат, не собирает детенышей в гнездо, а, наоборот, разбрасывает их. Она даже не перекусывает пуповину. Тем не менее, если собственный помет такой самки на несколько дней забрать от матери и подложить для выкармливания нормальной приемной матери, а самой мутантной самке подложить более старших по возрасту 4-дневных детенышей, то впоследствии у нее может проявиться материнское поведение. Для этого самку с детенышами следует поместить в очень тесную клетку, в которой она не может "раскидать" детенышей и поэтому оказывается в состоянии вырастить большую часть помета. При этом помимо кормления молоком самка способна вылизывать детенышей, однако ни строить гнездо, ни подтаскивать их к нему она не может. Было также показано, что при мутации sg изменены уровни лютеинизирующего и фолликулостимулирующего гормонов в плазме крови (Guastavino et al., 1992).
Плавание. При некоторых неврологических мутациях, затрагивающих внутреннее ухо и органы равновесия, эта способность полностью отсутствует, тогда как у мозжечковых мутантов плавание нарушено меньше, чем ходьба. Возможно, что этот тип локомоции, как более древний и более примитивный, базируется на более простых, а следовательно, менее затронутых анатомическими аномалиями сетях нейронов.
Исследовательское поведение. При мутации nervous (nv), повреждающей клетки Пуркинье мозжечка и его центральные ядра, исследовательское поведение изменяется незначительно. Число норковых реакций (заглядываний в отверстия, проделанные в полу камеры) не отличается от нормы. В то же время nv и другие мутанты характеризуются персеверацией реакций, т.е. повторным обследованием одних и тех участков пространства (отверстий-норок). По сравнению с нормальными животными у мутантов резко ослаблен процесс привыкания исследовательского поведения, иными словами, у них нарушен один из наиболее универсальных процессов адаптации к внешним условиям. Следует, однако, помнить, что причиной сильных аномалий исследовательского поведения этих мутантов может быть, по крайней мере частично, измененная моторика.
Реакция чередования. Попеременное перемещение то вправо, то влево при обследовании пространства, при поисковом поведении, при помещении в достаточно знакомую обстановку — характерная черта лабораторных мышей и крыс. У мутантов sg эта способности полностью отсутствует.
Аналогичный дефект отмечен у мутантов rl, wv и при мутации purkinje cell degeneration (pcd). Полагают, что данное нарушение есть следствие аномалий в анатомии и/или нейрохимии гиппокампа.
Обучение в лабиринте; пространственная память. Аномальное строение гиппокампа с нарушением расположения слоев при мутации Hippocampal lamination defect (Hid), свойственное линии BALB/с, выявляется только гистологически, а специфических изменений поведения, характерных для данной мутации, не описано. При мутации rl нарушаются некоторые виды поведения. При мутации Hid обучение реакции пассивного избегания не страдает, но оно резко изменено у rl, у которой были нарушены даже начальные этапы формирования навыка поиска пищи в радиальном лабиринте. Несмотря на фенотипическое сходство особенностей поведения мутантов rl и sg, аномалии поведения в радиальном лабиринте и в водном лабиринте Морриса были более сильными у rl.
Развитие генной инженерии и молекулярной биологии сделало возможным получение так называемых трансгенных животных, упоминавшихся в разделах 8.5.3.2 и 8.6.3.3. Напомним, что в геном животного (в настоящее время используются почти исключительно мыши) вводится новый генетический материал — это может быть или участок ДНК, кодирующий измененный ген, уже имеющийся у реципиента, или ген от животного другого вида, например крысы. Выделенный фрагмент ДНК вводится в геном реципиента (точнее, в его зародыш) разными способами. Во-первых, можно произвести прямую инъекцию ДНК в оплодотворенную яйцеклетку. Во-вторых, используя вектор ретровируса, можно ввести ДНК в развивающийся эмбрион, причем наиболее эффективно введение на стадии морулы. Третий способ — это введение в бластоцисту стволовых эмбриональных клеток (с измененным геномом).
Специальные молекулярно-биологические методы обеспечивают достаточно надежную экспрессию новой ДНК в геноме реципиента, иначе введенный в геном фрагмент может сохраняться в латентном виде, не обнаруживая себя (Crenshow et al., 1992).
Как и при работе с химерными животными, для успешного исследования трансгенного организма необходимо уметь четко идентифицировать, у кого именно из мышей данного помета генотип имеет новый, необычный ген. Для этого важно найти надежные маркеры — либо биохимические, либо молекулярно-биологические. Иногда носители такого нового гена внешне отличаются от нормальных собратьев.
Эксперименты по введению фрагмента ДНК с делецией по гену, кодирующему белок CREB, были упомянуты выше (см.: 8.6.3.3).
Интересные результаты дали исследования мышей с мутантным, т.е. дефектным геном, кодирующим дофаминовый рецептор D-1. Животные с таким дефектом развиваются с задержкой и погибают, когда прекращается материнское вскармливание. Однако они успешно выживают, если их перевести на искусственное вскармливание жидкой пищей. У этих животных вполне нормальная координация движений, хотя при исследовательском поведении у них меньше ''стоек", чем у нормальных животных тех же пометов. В нейронах стриатума у них нет связывания веществ-лигандов, специфичных для D-1 рецепторов, хотя нейроны, на которых такие рецепторы находятся в норме, у мышей-нокаутов есть и сохраняются вплоть до взрослого состояния.
Размер мозга у мышей-нокаутов был несколько меньше, однако общее анатомическое строение мозга не менялось. Особенности были отмечены в цитоархитектонике и нейрохимии стриатума, а также в некоторых физиологических реакциях. Каждый из двух структурных компонентов полосатого тела — стриосомы и матрикс (см.: Groves, 1983; White, 1989) — у мутантных мышей имел свои, отличные от нормы особенности. В матриксе наиболее заметным признаком было резкое снижение числа клеток, иммунореактивных к динорфину. В системе стриосом снижалось специфическое окрашивание, которое свидетельствовало об изменениях в содержании опиоидных пептидов. Наиболее интересной особенностью поведения мышей, лишенных рецепторов класса D-1, было отсутствие активирующего эффекта введения кокаина. Можно ожидать, что этот объект станет перспективной моделью для выяснения механизмов влияния наркотических веществ на мозг.
Анализ работы мозга и особенностей поведения трансгенных животных ставит перед исследователями вопрос, уже не раз поднимавшйся в связи с анализом эффекта мутаций. Мутация данного гена может влиять на мозг и поведение не только в связи с выключением его специфической функции, но и потому, что собственно формирование структуры мозга (в данном примере — стриатума) в процессе нейрогенеза у таких мышей также может происходить аномально. Возвращаясь к терминологии классической генетики, мы пока не знаем, в чем заключаются плейотропные эффекты "трансгенной" мутации.
Тем не менее, метод создания трансгенных животных дает ценнейшие данные о специфических функциях тех или иных генов, например в процессе развития мозга, поэтому список линий мышей с измененными генами, а также мышей-нокаутов быстро пополняется. В ряде случаев создание подобных моделей необходимо для практических целей, например для создания мышей без нуклеотидной последовательности, кодирующей так называемый прионный белок. Как известно, молекулы этого белка считаются инфекционным началом особой группы нейродегенеративных заболеваний, в том числе болезни овец — скрэпи, от которой ежегодно погибают тысячи животных. В связи с этим практическая задача состоит в создании овец-нокаутов. В качестве первого этапа этого исследования были получены мыши-нокауты по гену прионного белка. Они вполне жизнеспособны и внешне не отличаются от обычных. В пределах нормы сохраняется также обучение пространственному навыку — ориентации в водном лабиринте Морриса.
Были также получены мыши с полностью выключенной функцией гена Dvl1 (одного из трех генов мыши, гомологичных гену полярности сегментов дрозофилы). Животные оказались жизнеспособными, фертильными, у них не было морфологических отклонений в мозге, но отмечались некоторые особенности поведения и физиологии. Во-первых, это была аномалия организации сенсорного притока (gating) в слуховом и тактильном анализаторе. Во-вторых, неожиданно выяснилось, что для них было характерно снижение социальных контактов. Это произошло следующим образом. Отличительная особенность поведения мышей линии 129/SvSv — это "подстригание усов (вибрисс)" соседям по клетке, так что по достижении взрослого состояния значительный процент животных оказывается лишенным вибрисс (такое поведение описано у самцов и самок нескольких инбредных, линий мышей). Гомозиготные мыши-нокауты по упомянутому гену сохраняли свои усы в целости, что было причинно связано с резким уменьшением социальных контактов между этими животными. У них было нарушено и гнездостроительное поведение.
Особенности поведения (а также нейрохимической и морфологической организации мозга) мышей разных инбредных линий следует учитывать при оценке эффектов искусственных "генноинженерных" мутаций. Целый ряд досадных ошибок в выборе "фоновой" линии заставил специалистов, работающих в этой области, провести подробный анализ данных и выработать рекомендации по предпочтительному, наиболее информативному использованию той или иной линии при мутационном вмешательстве в какую-либо из систем мозга.
Аудиогенная эпилепсия. Судороги, развивающиеся при действии сильного звука у лабораторных мышей и крыс, рассматриваются как модель эпилепсии человека и интенсивно исследуются в плане оценки роли генотипа в генезе этого явления.
У мышей чувствительность к звуку была впервые отмечена д-ром Студенцовым, работавшим в лаборатории И.П.Павлова. Серьезное изучение этого феномена у мышей началось только с 1947 г., когда у мышей линии DBA/2 в определенный возрастной период (примерно с 15-го по 25-й дни жизни) обнаружили аудиогенные судороги, нередко с летальным исходом, связанным с остановкой дыхания.
В середине 40-х годов Л.В.Крушинский и Л.Н.Молодкина в МГУ путем селекции вывели линию белых лабораторных крыс, которые на сильный звук (около 100 децибелл над порогом слышимости человека) реагировали сначала мощным двигательным возбуждением, а затем клонико-тоническими и тоническими судорогами. Чувствительность к звуку имеют и беспородные крысы, и животные других линий (например, Wag, August), а также крысы популяции Вистар, однако среди них и двигательное возбуждение, и судороги встречаются, как правило, у небольшого процента животных. Крысы линии КМ, за редкими исключениями, обладают высокой чувствительностью к звуку. Известны также две другие линии чувствительных к звуку крыс: GERP (genetic epilepcy response prone), выведенные в Аризонском университете (США), которые различаются между собой по уровню этой чувствительности (Riegel et al., 1986).
Указанные линии крыс и мышей интенсивно исследуются как нейрохимиками, так и фармакологами. Установлено, что у чувствительных к звуку животных показатели состояния и ГАМК-ергической системы, и моноаминергических систем достоверно отличаются от нечувствительных. В эту патологию вовлечена также и холинергическая система мозга.
Данные литературных источников о генетическом определении чувствительности к звуку у мышей несколько различаются. В одной из наиболее известных работ американских исследователей Дж.Фуллера и Р.Коллинза (Collins, Fuller, 1968) по скрещиванию чувствительной и нечувствительной к звуку линий — DBA/2 и C57BL/6J соответственно — было показано, что этот признак определяется одним геном, локализованным на 12-й хромосоме и получившим название asp (audiogenic seizure prone}. В настоящее время он называется asp-1. Вместе с тем данные, полученные на батарее рекомбинантных инбредных линий (см.: 8.6.6.3) с использованием чувствительной и нечувствительной родительских линий, показали, что признак детерминирован более чем одним геном. Второй ген, ответственный за этот признак, — asp-2 — был локализован на 4-й хромосоме.
Изучение характера наследования повышенной чувствительности к звуку у крыс методом диаллельного скрещивания, который описан в разделе 8.6.6.2, показало, что данный признак имеет полигенную природу, действие генов аддитивно и гены, детерминирующие нечувствительность к звуку, доминируют. Чувствительность к звуку, характерная для крыс линии КМ, является почти полностью рецессивным признаком (Романова и др., 1975). В природных популяциях и в лабораторных линиях практически не встречаются животные, столь чувствительные к звуку, как крысы линии КМ. Вопрос о биологической роли аномальной чувствительности к звуку, точнее о его эволюционном генезе, не только не исследован, но практически никогда не поднимается.
У крыс и мышей, предрасположенных к аудиогенным припадкам, найдены также эндокринные расстройства, нарушения водно-солевого обмена и метаболизма пиридоксина (Loscher, 1992). Показано, что у линии GERP-9 (см. выше) повышено количество ГАМК-ергических нейронов заднего двухолмия, причем, в отличие от резистентных к звуку животных, у чувствительных увеличено число мелких нейронов, а их средние размеры также меньше, чем у нечувствительных крыс. Эти нейроны аномальны по строению сомы и отростков, а количество их положительно коррелирует с тяжестью аудиогенного приступа (Daily et al., 1989).
Молекулярно-генетические исследования у грызунов с аудиогенными припадками немногочисленны. Недавно было показано, что у крыс линии КМ отсутствуют определенные гипервариабельные фрагменты ДНК (маркеры, свойственные резистентым лабораторным животным), которые, вероятно, отражают структурные изменения в геноме и связаны с проявлением аудиогенного приступа. Кроме того, у крыс этой линии сразу после припадка подавлена экспрессия быстрых генов раннего действия.
Аудиогенной чувствительности у мышей и крыс разных генотипов сопутствуют нарушения связывания и измененный синтез как возбуждающих аминокислот и ГАМК, так и изменение содержания (как правило, снижение) катехоламинов мозга (Loscher, 1992). У таких животных изменены мембранные процессы в нейронах и/или в глиальных клетках, у них имеются также региональные особенности морфологии мозга. Характер генетической архитектуры признака также неодинаков у разных линий (см. выше).
Признак "чувствительность к звуку" используется для исследования механизма развития судорожных состояний. Линии мышей и крыс, обладающие аудиогенной чувствительностью, служат моделями эпилепсии человека. Аудиогенная эпилепсия относится к категории патологических состояний, которые провоцируются внешней стимуляцией, иначе говоря рефлекторно. К этой группе патологии относится также так называемая "фотоэпилепсия" человека и некоторых животных и ряд других состояний (см. ниже). У человека практически не бывает аудиогенных судорог в том виде, в каком они обнаруживаются у грызунов. Описаны, однако, эпилептические припадки человека, которые провоцируются музыкой (Wieser et al., 1997). Это означает, что аудиогенная эпилепсия грызунов как модель заболевания человека имеет определенные недостатки. В тоже время легкость провокации судорог, четкость и воспроизводимость феномена, а также возможность проведения межлинейных сравнений на физиологическом, биохимическом и молекулярном уровнях делают исследование этого признака чрезвычайно популярным.
Другие виды рефлекторной эпилепсии. Генетически детерминированные судороги, возникающие в ответ на те или иные сенсорные воздействия, т.е. рефлекторно, описаны и у других видов животных.
Одна из сенегальских популяций павианов (Рарio papio) насчитывает значительное число особей, у которых мелькающий свет провоцирует миоклонические судороги. Эти животные представляют собой единственную генетическую модель эпилепсии на приматах, поэтому ее значение трудно переоценить (Loscher, 1992).
У кур описана аутосомная рецессивная мутация, при которой различные сенсорные стимулы вызывают тонико-клонические судороги. Наиболее четко этот феномен проявляется при использовании мелькающего света. Интересно отметить, что у кур, как и у мышей и крыс, часть рефлекторного судорожного приступа заключается в интенсивной локомоции. Этот феномен специфичен для данных видов животных и для данного типа судорог, поскольку практически не встречается при химически вызванных судорогах и при судорогах, развивающихся в ответ на электрошок.
У монгольских песчанок (Meriones unguiculatus), содержавшихся в университете штата Техас, были обнаружены животные (примерно 27%), у которых судороги развивались в ответ на новую обстановку. В дальнейшем оказалось, что судороги развиваются в ответ на такие воздействия, как включение света и звука, потряхивание клетки и даже при взятии зверьков в руки. Частота их возникновения была выше у животных, полученных в результате специальной селекции, однако, как и при аудиогенной эпилепсии крыс и мышей, внешние воздействия могли сильно изменять их проявление. В настоящее время разработана методика провокации таких судорог дозированной струей воздуха, направленной на голову животного. Это позволило добиться достаточно стабильного проявления судорог и успешно использовать модель для тестирования антиэпилептических препаратов (Ldscher, 1992).
Спонтанно возникающие судороги. Эпилепсия собак как хроническое заболевание, проявляющееся в частых спонтанных судорогах, напоминает аналогичную болезнь человека и поэтому привлекает значительное внимание исследователей. Спонтанные эпилептиформные судороги чаще встречаются у миниатюрных шнауцеров, бассетов, колли и коккер-спаниелей. Однако наиболее подвержены эпилепсии бигли, которые в связи с этим нередко используются в лабораториях. Этому способствует сходство и в развитии припадков у человека и собаки, и в характере влияния на них известных противосудорожных препаратов. В ряде случаев, однако, судороги, развивающиеся у собак, нельзя назвать спонтанными, поскольку они чаще всего случаются в состоянии сильного эмоционального возбуждения. Только у собак обнаружены так называемые "большие припадки" с клонально-тоническими судорогами, сходные с таковыми человека, что делает данную модель особенно ценной.
В одной из лабораторий города Страсбурга (Франция) было обнаружено, что у 30% крыс из популяции Вистар, получившей название Wag/Rii, возникали спонтанные судорожные состояния, более всего напоминавшие малые припадки, характерные для эпилепсии человека. Электрофизиологическое исследование подтвердило, что эти приступы действительно имеют эпилептическую природу (так называемые "абсансы"). Они остаются практически неохарактеризованными генетически, видимо, вследствие той же особенности признака — чрезвычайной зависимости от внешних условий, которая определяет их высокую вариабельность (Loscher, 1992).
Как говорилось выше, мыши с неврологической мутацией tottering, локализованной в 8-й хромосоме, также характеризуются спонтанно возникающими судорогами и типичными для эпилепсии изменениями ЭЭГ. Для этой мутации характерны изменения в норадренергической иннервации ряда отделов мозга. Это моногенный дефект, при котором теоретически возможно проследить генез мозговых аномалий, вызывающих судороги. Участие норадренергической системы в генезе патологической картины при этой мутации, возможно, опосредовано системой гликогенолиза. Вследствие нарушения обмена углеводов у мышей этой линии формируется избыточное число мозговых глюкорецепторов, вслед за чем изменяется и число адренорецепторов.
Нарколепсия. Это заболевание проявляется у человека (с частотой 0,02—0,09% в большинстве обследованных популяций) в виде аномальной сонливости в дневные часы, нарушений фазы ночного сна с быстрыми движениями глаз (БДГ-сон) и катаплексии (резкого падения мышечного тонуса в ответ на средовые стимулы).
В начале 70-х годов две группы исследователей независимо описали подобную аномалию у собак — пуделей и биглей. В их потомстве, однако, заболевание не проявлялось. Позднее этот же признак, четко передающийся генетически, был описан у доберман-пинчеров и лабрадоров. Такие линии собак в настоящее время поддерживаются и исследуются как модель заболевания человека. В указанных линиях собак нарколепсия передается как аугосомный рецессивный признак с полной пенетрантностью (Mignot et al., 1992).
Электроэнцефалографические обследования собак-нарколептиков выявили у них нарушения характерной для сна электрической активности, которые свойственны нарколепсии человека. Для мутантных особей наиболее характерными были приступы катаплексии, причем они провоцировались главным образом эмоционально положительными стимулами — перспективой игры, получением пищи, половым возбуждением. Приступ представляет собой быстро развивающуюся потерю тонуса мускулатуры, поддерживающей позу, при полном выключении моно- и полисинаптических спинномозговых рефлексов. Помимо катаплексии у собак этих линий иногда наступали короткие эпизоды дневного сна, также индуцированные предъявлением пищи.
Несмотря на значительную индивидуальную изменчивость тяжести симптомов, возраста начала заболевания, его течения, в картине нарколепсии выявляются и межпородные различия. У пуделей она начинается в более позднем возрасте и протекает тяжелее, чем у доберманов и лабрадоров. Авторы полагают, что разнообразие клинических проявлений нарколепсии собак определяется сложным характером нарушений, лежащих в ее основе.
Нейрофизиологическое исследование мозга собак с нарколепсией показало, что в регуляции катаплексических приступов участвует по меньшей мере две мозговые системы — холинергичсская система моста (интерпедункулярное ядро — ретикулярные ядра моста) и система нейронов голубого пятна, проецирующихся на нейроны моста и миндалевидного комплекса. Нейроны миндалины в свою очередь проецируются в ствол мозга (голубое пятно и /или холинергические системы ствола), что может объяснять роль эмоционального возбуждения в запуске механизма катаплексического приступа.
У человека проявления нарколепсии ассоциированы с экспрессией антигена HLA-DR2, связанного с функцией главного локуса гистосовместимости. В отличие от человека, у собак-нарколептиков не обнаружилось такого связанного с локусом Н-2 белка, присутствие которого коррелировало бы с наличием заболевания.
Аутосомно-рецессивный ген, определяющий наследственную катаплексию у собак, был назван "саnаrс-1'' Дальнейшие исследования показали, что ответственные за нарколептические симптомы локусы и у человека, и у собаки имеют четкую связь с генами иммунной системы. Можно предположить,что существует связь этого заболевания с аномалиями в иммунной сфере.
Кататония — это синдром, характеризующийся застыванием в разных позах или стереотипным двигательным возбуждением. Этот синдром считается признаком шизофрении, хотя встречается и при других психических заболеваниях. Применительно к животным термин впервые использовали для обозначения неспецифической реакции на некоторые воздействия, в частности фармакологические, однако сам феномен был известен значительно раньше под названием "животный гипноз".
Кататонический синдром имеет два полюса — гиперкинетический (стереотипное, нецеленаправленное возбуждение) и акинетический (ступор). В последнем случае может обнаруживаться восковая гибкость (каталепсия), или мышечное напряжение. Собственно "животный гипноз" и представляет собой состояние ступора.
Каталепсия у крыс. Генетические, нейрофизиологические и нейрохимические аспекты кататонии у животных подробно освещаются в монографии новосибирского ученого В.Г. Колпакова (1990; см. также: Попова и др., 1978). В частности, под его руководством была проведена селекция лабораторных крыс на высокую предрасположенность к формированию каталептической реакции (т.е. застыванию в неподвижной позе) при приподнимании за передние лапы. Эффект селекции (усиление выраженности признака) был получен за 2 поколения, что, как мы уже видели раньше в случае селекции Phormia regina (см.: 8.4.2), свидетельствует об участии в определении этого признака небольшого числа генов. В то же время доля животных с таким признаком в селектируемой линии увеличивалась достаточно медленно и лишь к 21-му поколению селекции достигла 90%. По-видимому, экспрессия признака находится под сложным генетическим контролем, в котором участвует значительное число генов-модификаторов. У этих крыс отмечены отклонения в функции дофаминергической системы мозга.
Животные, у которых подобная каталептическая реакция проявляется при минимальном внешнем воздействии (при приподнимании их передних конечностей с помощью простого стержня), обнаруживаются практически во всех тестированных популяциях лабораторных крыс. Более того, попытка автора провести селекцию на отсутствие предрасположенности к каталепсии не увенчалась успехом.
Эти экспериментальные факты могут свидетельствовать о том, что склонность к формированию подобных реакций находится в диапазоне нормальных и жизненно важных физиологических ответов организма, составляя филогенетически древнюю часть репертуара специфических механизмов адаптации. Представляет большой интерес возможная связь между генетическими, физиологическими и нейрохимическими основами этих реакций и механизмами генеза патологии человека, в частности патогенеза шизофрении.
Каталепсия у мышей. Каталептическое состояние, которое развивается рефлекторно в ответ на тактильное раздражение кожи загривка, описано у мышей линии СBA (Куликов, Козлачкова, 1989) (так называемая "щипковая" кататония). По данным скрещивания линий СВА и AKR, этот признак имеет однолокусное рецессивное наследование и аутосомную локализацию. Анализ нейрохимических показателей у мышей СВА и их гибридов позволяет предположить, что данная аномалия связана с особенностями функции серотонинергической системы стриатума, тогда как дофаминергическая система у них практически не нарушена, что не совпадает с данными, которые были получены на крысах, предрасположенных к кататонии.
Физиологические основы. Строго говоря, состояние, именуемое стрессом, не относится к числу патологических реакций организма, однако формирование физиологических реакций на неблагоприятные воздействия, которое изучается генетическими методами, как правило, рассматривается в рамках моделей тех или иных патологических состояний человека. В связи с этим мы сочли возможным поместить данные по экспериментальному исследованию генетического контроля стресс-реакций в этом разделе.
Способность организма противостоять и адекватно реагировать на внешние и внутренние стимулы в условиях действия пугающих или вредоносных агентов (так называемых ''стрессирующих агентов") составляет суть процесса успешной адаптации к изменяющимся условиям среды. При действии стрессоров в организме запускается ряд нейрохимических и нейроэндокринных процессов, а также облегчаются некоторые поведенческие реакции. На основе способности к обучению и элементарной рассудочной деятельности, ориентации в пространстве и прочего животное оптимизирует поведение с целью избегнуть вредоносного влияния. Очевидно, что при этом обнаруживается генетическая изменчивость поведения и физиологических реакций.
Практически все экспериментальные исследования поведения в связи со стресс-реактивностью — это работы, в которых анализируется достаточно быстрая физиологическая реакция организма на изменение внешней обстановки (помещение в "открытое поле") и/или на болевое воздействие.
Обычно предполагается, что такие воздействия бывают "умеренно стрессирующими". Более сильные воздействия — это иммобилизация, эфирный наркоз, удары тока, в особенности повторяющиеся, избежать которых животное не может. Временная шкала, в которой развиваются подобиые ответные реакции, — это десятки секунд или минуты, Такие реакции представляют собой "нормальный" физиологический процесс, в отличие от классического адаптационного синдрома в том виде, в каком он был описан его первооткрывателем Г,Селье (1960; 1972), В связи с этим вопрос о возможном параллелизме в закономерностях развития физиологически нормальной или "естественной" стресс-реакции и патологического процесса "стресса" с его фазами активации, стабилизации и истощения еще надлежит внимательно проанализировать, Очевидно, что "физиологический" стресс встречается у животных в обычных условиях, тогда как "патологический" стресс — порождение физиологического эксперимента, в котором исследователь стремится воспроизвести на животных условия, воздействующие на человека в его часто аномальной среде обитания. Некоторой аналогией стресса в понимании Селье, по нашему мнению, являются аномальные состояния, которые развиваются у крыс при содержании их в замкнутом пространстве при неограниченном размножении и перенаселении (знаменитая работа Дж. Колхауна, Calhoun, 1962; см: 5.5.1). Следует подчеркнуть, что эти явления характерны не для естественной, а для искусственной среды обитания.
Первым и наиболее важным физиологическим ответом на действие стрессирующего фактора является активация оси "гипоталамус—гипофиз—надпочечники". На основании современных знаний представляется следующий путь такой активации: неблагоприятные для организма стимулы ведут к активации паравентрикулярных ядер гипоталамуса, вследствие чего нейроны этой структуры выделяют кортикотропин-рилизинг-фактор — кортиколиберин. Кортиколиберин попадает в воротную сосудистую систему, орошающую переднюю долю гипофиза. Ответом передней доли гипофиза является секреция бета-эндорфинов и АКТГ, причем последний запускает в коре надпочечников реакцию секреции глюкокортикоидов. Глюкокортикоиды (это кортикостерон у мыши и крысы, кортизол — у человека, обезьян и собаки), помимо выполнения своих основных физиологических функций, участвуют также в регуляции уровня АКТГ в крови, воздействуя по механизму отрицательной обратной связи на гипофиз, а также на структуры гипоталамического уровня (и структуры, лежащие более "высоко"), т.е. на функцию всех участков оси "гипоталамус-гипофиз-надпочечники". Воздействие на организм стрессового фактора активирует, к тому же, и симпатическую нервную систему, в результате чего и в ЦНС, и на периферии выделяются адреналин и норадреналин.
Исследование процессов, сопровождающих активацию данной "оси", можно проводить на метаболическом уровне, а также на уровне поведения животного. Рост уровня глюкокортикоидов усиливает использование глюкозы, липолиз, глюконеогенез и снижает уровень белкового синтеза.
Рост концентрации катехоламинов увеличивает частоту биения сердца и повышает давление крови, что способствует улучшению питания тканей и снабжения их кислородом. Физиологический ответ на стрессор выражается и в общей активации поведения. При этом можно наблюдать одну из трех видов реакций: драку, бегство или реакцию застывания или затаивания (freezing).
Статается, что каждое из этих состояний связано с преимущественным вовлечением разных нервных структур, а также разных гормональных систем.
Физиологические, эндокринные и нейрохимические особенности реакций на умеренные по силе стрессоры и их генетический контроль достаточно успешно исследуются в опытах на селектированных и инбредных линиях лабораторных грызунов (см.: Gray, 1987 Полетаева, Романова, 1990; Driscoll, 1992; см. также: 8.6.3.2). В генетических исследованиях поведения считается очень продуктивным анализ и эмоциональной реактивности животных.
В первой работе, связанной с применением генетического подхода к исследованию эмоциональной реактивности животных, английский исследователь П.Бродхерст (1975) показал, что этот признак в большой части случаев находится в обратной зависимости от уровня двигательной активности животных, однако это соотношение далеко не всегда выполняется (в частности, в связи с тем, что у некоторых животных состояние страха вызывает не подавление активности, а бегство). Более того, физиологические исследования показали, что количество реакций дефекации (а значит, и эмоциональная реактивность) более четко коррелирует с повышением уровня кортикостерона в плазме крови (т.е. с показателями стресс-реакции), чем уровень двигательной активности.
Римские линии крыс (RHA, RL4). Как упоминалось выше (см.: 8.6.3.2), эти линии были селектированы на быстрое и медленное формирование реакции активного избегания болевого стимула — удара электрического тока через пол в челночной камере. В физиологических показателях и поведении крыс RHA и RLA были обнаружены многочисленные различия.
Напомним, что данные этологического анализа реакции активного избегания позволяют предполагать, что линия RLA отвечает на болевой стимул реакцией застывания (что делает их иплохо избегающими"), a RHA — хорошо избегают удара тока, так как для них характерна реакция на боль в виде бегства.
Вызванное состоянием умеренного стресса повышение в плазме крови уровней кортикостерона, АКТГ и пролактина, а также количества актов дефекации были выше у линии RLA со слабой способностью к избеганию тока. В то же время при действии сильных стрессоров, таких как эфирный наркоз, удар тока и иммобилизация, межлинейные различия у "римских" крыс были выражены слабо.
У линии RLA фоновый уровень АКТГ был ниже, а кортикостерона выше, чем у RHA.
Таким образом, по крайней мере частично межлинейные различия в поведении можно объяснить различиями в гормональной активности оси гипофиз-надпочечники, возникшими как ответ на отбор. Интересно отметить, что детеныши крыс этих линий не различаются по гормональному фону и реакциям на стрессоры вплоть до прекращения материнского вскармливания, т.е. до возраста 21 день (Walker et al., 1992).
Maudsley Reactive (MR), Maudsley Nonreactive (MNR) линии крыс (см. также: 8.6.3.2), селектированные на высокий и низкий уровни эмоциональности, мало изучены в аспекте развития у них реакций на стрессоры. Известно, однако, что у MR фоновый уровень давления крови и частоты сердечных сокращений выше, чем у MNR, а в ответ на зрительные и слуховые стимулы у крыс MR учащение пульса сильнее, чем у MNR.
У сиракузских линий крыс (SHA и SLA) (см.: 8.6.3.2), которые, различаясь по скорости формирования реакции избегания, не различаются по уровню двигательной активности, эмоциональная реактивность в "открытом поле" была выше у SLA, т.е. у плохо обучающихся крыс. У этой же линии был ниже, чем у SHA, уровень глюкокортикоидов в плазме и выше вес надпочечников. Таким образом, у римских и сиракузских пар линий, селектированных на разный уровень реакции избегания тока, а косвенно — на разный тип реакции на боль, корреляции с другими признаками поведения — двигательной активностью и эмоциональной реактивностью — оказались разными. Это заставляет относиться к данным по корреляции признаков поведения с большой осторожностью.
Исследование стресс-реактивности линий КНА и KLA (см.: 8.6.3.2), также созданных путем искусственного отбора на высокую и низкую способность к обучению реакции активного избегания, позволило выявить принципиально разные способы реагирования (две стратегии поведения) этих животных на стрессор (Жуков, 1997).
Генетические исследования социального стресса на мышах. Популяции диких мышей состоят из замкнутых "демов" — небольших групп особей, в состав которых входят доминант, несколько самок и подчиненных самцов. Попадание на территорию дема чужака вызывает агрессивную реакцию доминанта (см.: 5.5.1).
Роль генотипа в формировании социального доминирования была впервые исследована в работе Дж.Маклерна и Дж.Дефриза в 1972 г. (см.: McClearn, DeFries, 1972). Их опыты по созданию искусственных популяций лабораторных мышей разных генотипов показали, что и доминантное положение самца, и его последующий успех в размножении четко зависят от генотипа (см.: Эрман и Парсоне, 1984). Самцы линий А и BALB/c чаще занимали доминирующее положение, линия C57BL/6J — промежуточное, а самцы DBA/2J наиболее часто были подчиненными особями. Доминантный самец оказывался отцом 90% всех потомков.
Сходные данные были получены в работах Е.В.Науменко и его сотрудниками в Институте цитологии и генетики СОАН СССР в Новосибирске, проанализировавших состояние оси "гипоталамус-гипофиз—надпочечники" при объединении самцов-мышей разных линий в искусственные группы-популяции. Помимо того что способность к победе в агрессивных столкновениях оказалась различной у самцов исследованных линий, она была разной и в ходе установления доминантно-субординантных отношений. Когда они начинают активно формироваться, то у самцов, которые в дальнейшем будут доминантами, эта активность бывает повышенной. Сустановлением иерархии более высокая активность системы бывает у подчиненных особей (Науменко и др., 1983; Серова, Науменко, 1990). Изменения уровня кортикостерона в условиях хронического стресса у мышей, потерпевших поражения в столкновениях, также зависели от генотипа. На основании анализа межлинейных различий и особенностей показателей гормонального статуса у мышей-гибридов можно предполагать, что системы генетического контроля содержания кортикостерона в плазме в условиях физиологической нормы и при стрессе независимы друг от друга.
Данные Е.В.Науменко и его коллег получены на мышах-самцах, которые содержались в искусственных группах-популяциях. Встает вопрос, в какой степени процессы, выявленные в таких условиях у лабораторных мышей, помогают пониманию роли генотипа в особенностях социального поведения дикой формы домовой мыши? Считается, что численность грызунов в природе регулируется рядом факторов, причем важная роль при этом принадлежит внутрипопуляционным отношениям (Дэвис, Кристиан, 1977; Шилов, 1977; см. также: Дьюсбери, 1981; Эрман, Парсонс, 1984). Они, в свою очередь, очевидным образом связаны с плотностью популяции. Как следует из вышеизложенного, генотип относится к одному из факторов, определяющих особенности стресс-реакции на перенаселение.
В исследованиях механизмов алкогольной интоксикации, начатых в 60-е годы XX в., разрабатывались разнообразные подходы, важнейшей частью которых было создание экспериментальных моделей алкогольной мотивации. Так, например, лишение воды и/или пищи, стрессовые воздействия, хирургические и фармакологические вмешательства должны были имитировать условия, при которых алкоголизм формируется у человека. Однако такие модели не были достаточно эффективными. Основное требование к экспериментальной модели алкоголизма заключается в том, что животное должно добровольно пить значимое количество алкоголя (этанола) в условиях свободного выбора.
Существуют методы отбора таких животных из генетически гетерогенных популяций мышей или крыс (из беспородных лабораторных животных, крыс разных стоков или гибридных мышей), после чего их можно исследовать в специальных экспериментах. В подобных случаях говорят о фенотипической модели алкоголизма. В то же время отобранные таким образом животные могут служить исходной популяцией для последующей селекции на высокие и низкие значения признаков, связанных с формированием алкоголизма. Таким путем создаются генетические модели. Еще одним вариантом генетической модели алкоголизма может быть выявление различий в поведении, связанном с приемом алкоголя, между мышами инбредных линий, что было обнаружено на практике при сравнении линий C57BL/6J и DBA/2J (см. ниже).
Модель алкогольной мотивации, в частности генетическая, должна удовлетворять двум важным условиям. Во-первых, такие животные должны, как уже упоминалось, добровольно пить такие дозы алкоголя, которые небезразличны для обмена веществ (для крыс и мышей — это более 5 г/кг в сутки), а во-вторых, в условиях временного ограничения доступа к алкоголю у них можно оценить стремление к его потреблению. Важным феноменом в экспериментальном исследовании алкоголизма является так называемый положительный алкоголь-депривационный эффект (Салимов, 1998). Данный эффект выражается в увеличении потребления алкоголя в первые 1—2 часа после кратковременного лишения животного доступа к нему.
В селекционных экспериментах, проведенных в разные годы, было создано нескольких пар линий крыс, отличающихся по количеству потребляемого этанола (Crabbe et al., 1993, 1996; Feller, Crabbe, 1991). Это линии UCh-B/UCh-A (Чили), Alco Alcohol/Alko Non-Alcohol (AA/ANA, Хельсинки), Preferring/Non-Preferring (P/NP, США), High Alcohol Drinking/Low Alcohol Drinking (HAD/LAD, США) и High, Control, Low Alcohol Sensitive (HAS, CASt LAS, США).
Об успешности такой селекции говорит тот факт, что крысы линии АА в 55-м поколении селекции потребляли примерно 7,6 г/кг этанола в день, тогда как у ANA эта величина достигала всего 0,4 г/кг. У крыс линии АА легко формировалась реакция нажатия на рычаг для получения спирта, тогда как у крыс ANA этого сделать было нельзя, поскольку для них этанол не был подкрепляющим агентом. Сам факт создания таких линий доказывает наличие генетического контроля в формировании пристрастия к алкоголю, т.е. алкогольной мотивации. Мотивация удовлетворяется при приеме алкоголя. Иными словами, в терминах физиологии обучения, алкоголь обладает свойствами подкрепляющего агента (Sinclair, 1992).
Упоминавшийся выше положительный алкоголь-депривационный эффект в сущности является частным случаем психологического феномена положительного контраста, т.е. проявлением ожидания положительного подкрепления. Он формируется в результате ассоциации центральных эффектов алкоголя и его свойств как раздражителя с активацией системы мозговых структур, связанных с положительным подкреплением. В основе алкоголь-депривационного эффекта лежит ожидание ранее уже испытанного удовольствия от приема алкоголя. Если же данный стимул не приобрел свойств положительного подкрепления, то ограничение доступа к нему не повлияет на стремление животного к его получению.
Сложность экспериментального генетического исследования алкоголизма заключается в том, что помимо формирования такой специфической мотивации это заболевание характеризуется целым набором специфических признаков, которые все вместе до сих пор не удалось воспроизвести ни в одной модели. Практически все существующие генетические модели алкоголизма воспроизводят одну, две или больше (реже) из этих патологий.
У генетических линий животных, селектированных на высокую и низкую интенсивность стремления к употреблению алкоголя, исследуют как отдельные симптомы алкоголизма (см.: ниже), так и более "интегральный уровень" — состояние многих физиологических систем. Примером исследования второй группы является работа P.M. Салимова (1998), в которой, в частности, сопоставлялся общий характер особенностей поведения крыс нескольких "алкогольных" линий с особенностями протекания болезни у человека.
Интегральный уровень экспериментального исследования алкоголизма. Исследования последних десятилетий говорят о существовании типологических различий в клинических проявлениях зависимости от этанола у человека (Салимов, 1998). В целом считается установленным наличие двух категорий индивидов. Один из признаков, по которым они различаются — это бо'льшая (тип 1) или меньшая (тип 2) зависимость алкогольной мотивации от провоцирующего влияния внешних условий, а также некоторые особенности поведения таких людей. В частности, алкоголики 1-го типа мало интересуются новизной, обнаруживают повышенную тревожность, пассивность и пессимизм, тогда как больные, которые относятся ко 2-му типу, характеризуются высоким стремлением к новизне, решительностью и оптимизмом. Больные 1-го типа более восприимчивы к попыткам психотерапевтического воздействия, а индивидуумы 2-го типа демонстрируют навязчиво-устойчивую алкогольную мотивацию. Для лечения больных алкоголизмом 1-го типа в целом эффективно использование транквилизаторов и антидепрессантов, тогда как для больных 2-го типа эти вещества малоэффективны.
Подробные исследования нескольких пар линий крыс, селектированных на предпочтение растворов этанола, показали, что характер потребления спирта крысами линий HAD1 и HAD2, выведенными на основе стока N/Ni, в большой степени напоминают особенности алкоголиков 1-го типа. От "парных" им линий LAD1 и LAD2 их отличают не только достоверно большие количества потребляемого спирта, но и некоторые особенности поведения, иные, чем у крыс других "алкогольных" линий. Сходство с алкоголизмом 1-го типа заключается в том, что потребление алкоголя у таких животных изначально невысокое и нестабильное, их поведение характеризуется осторожностью, а в тесте "вынужденного плавания" — пассивностью.
Линии Р и АА (контрастные линиям NP и ANA), выведенные на основе популяции Вистар, особенностями потребления спирта в большей степени напоминают алкоголиков 2-го типа. Их характеризует высокий и очень устойчивый начальный уровень потребления спирта, они более активны в тесте "вынужденного плавания" и тревожность выражена незначительно. В тестах на исследовательское поведение и на способность активно избегать опасность крысы АА и Р, в сопоставлении с линиями HAD1 и HAD2, отличались не только от "парных" им не пьющих спирт линий, но и друг от друга. Таким образом, с большой долей уверенности можно говорить и о существовании различной генетической основы двух основных вариантов течения алкоголизма у человека.
Экспериментальное исследование отдельных симптомов алкоголизма. Другим подходом к экспериментальному исследованию генетических основ алкоголизма является подробное сравнение селектированных линий по особым чертам их поведения и последующий генетический анализ таких различий. В первой работе такого рода (см.: MacClern, DeFries, 1972) были установлены достоверные межлинейные различия в предпочтении растворов этанола мышами линий C57BL/6J и DBA/2J. Это послужило основанием для проведения дальнейших селекционных экспериментов.
В совокупности было создано более двадцати пар линий мышей и крыс, контрастных по поведению, связанному с потреблением алкоголя или с реакциями на его введение. Имеющиеся инбредные и селектированные линии позволяют исследовать отдельные группы признаков или симптомы алкоголизма. Таким образом, сложная картина симптомов алкоголизма в ряде генетических моделей может быть "расчленена" на более простые единицы. Это дает возможность оценивать роль фармакологических агентов в коррекции отдельных аномалий, связанных с приемом алкоголя.
Попытки селекции линий мышей и крыс, достоверно различающихся по степени предпочтения алкоголя, были в большинстве случаев успешными. Существование генетической основы в предрасположенности к развитию алкоголизма можно было считать доказанным. Следующей задачей стало выявление и картирование генетических элементов, определяющих сложный комплекс аномального поведения при алкоголизме: влечение, абстиненция, токсические реакции.
Ниже мы охарактеризуем некоторые из наиболее популярных генетических моделей, использующихся для исследования разных симптомов алкоголизма и для картирования ответственных за них генов.
Потребление больших количеств этанола, чувствительность к нему, и предпочтение алкоголя. Различия в предпочтении этанола инбредными линиями мышей C57BL/6J и DBA/2J6 были обнаружены в 1959 году в работе Дж. Маклерна и его коллег (см.: McClern, DeFries, 1972). Эти линии были использованы для создания рекомбинантных инбредных линий (см.: 8.6.6.3), т.е. группы линий, в которых аллели C57BL и DBA были перекомбинированы. Их изучение в соединении с методом картирования QTL (локусов количественных признаков, см.: 8.6.6.4) позволило в настоящее время картировать три гена, которые участвуют соответственно в формировании толерантности к этанолу (хромосома 12), его предпочтении безалкогольным растворам (хромосома 1) и в развитии гипногенного эффекта этанола (хромосома 7).
Толерантность этанола и его токсические эффекты. Эти физиологические признаки также исследуются на специфических генетических моделях. В качестве показателей токсичности этанола используют, например, длительность его гипногенного эффекта, т.е. сна после приема определенной дозы этанола. В лаборатории американского исследователя Дж. Крэба были выведены линии мышей LS (long sleep) и SS (short sleep) (Crabbe et al., 1993; 1996), используемые в решении множественных проблем алкоголизма. С помощью метода QTL исследуется батарея рекомбинантных инбредных линий, полученных на основе скрещивания LS и SS. Это позволяет локализовать гены, связанные с физиологическими различиями в реакции на этанол у мышей данных линий. Показано, что, по крайней мере частично, эти различия связаны с нейротензинергической системой мозга.
Мыши двух пар селектированных линий (FAST и SLOW), созданных путем отбора на различия в токсических эффектах этанола, т.е. на высокий и низкий уровни локомоции после его приема, отличались друг от друга и по уровню алкогольной мотивации, тестированной разными методами.
Устойчивость и чувствительность к гипотермическому эффекту после инъекции этанола также послужили контрастными признаками для селекции мышей. Выведенные линии НОТ и COLD достоверно различались по реакции на спирты с разной длиной молекулы и по чувствительности к большому спектру седативных препаратов (Crabbe et al., 1996).
Синдром отмены (приема алкоголя). Если животных, содержавшихся в атмосфере с высокой концентрацией паров этанола, помещали в обычные условия, когда попадание в организм этанола резко сокращалось, это вызывало различные патологические состояния, в частности вело к появлению судорог. В двух последовательно проведенных селекционных экспериментах было выведено по 2 пары линий мышей (Withdrawal Seizure Prone — WSP, и Resistant — WSR). В обоих случаях у мышей WSP отмена алкоголя вызывала появление судорог при взятии в руки. Мыши другой линии были устойчивы к отмене алкоголя. Генетический анализ этого феномена у межлинейных гибридов показал доминирование признака устойчивости к отмене алкоголя (Crabbe, Phillips, 1993).
Признаки, по которым можно было судить о чувствительности мышей указанных линий к этанолу и о толерантности к нему, у рекомбинантных линий распределялись по разному. Это означает, чгго явления, возникающие при отмене приема алкоголя, как и при его прямом влиянии на организм, связаны с разными системами генов.
При исследовании линий мышей и крыс, селектированных на разные признаки, связанные с действием алкоголя на организм, используются как гибридологический анализ и рекомбинантные инбредные линии, так и современные молекулярнобиологаческие подходы (метод картирования QTL).
Физиологическое изучение особенностей животных линий, контрастных по приверженности к приему спирта, по выраженности его токсических эффектов и синдрома отмены, дает импульс к поиску соответствующей патологии у человека. Поскольку существует возможность прямого картирования хотя бы части генов мыши, связанных с симптомами алкоголизма, это дает основание проводить прямое сопоставление с человеком. Как известно, в строении генома человека и мыши существует известный параллелизм (так называемая синтения), поэтому исследование генетических моделей алкоголизма на животных является залогом успехов в изучении предрасположенности человека к алкоголизму, а также его успешной профилактики и лечения.
Болезнь Альцгеймера — одна из наиболее распространенных форм старческого слабоумия, сопровождающаяся специфической картиной нейродегенеративных явлений. Заболевание характеризуется отложением так называемого амилоидного белка в виде бляшек на аксонах нервных клеток и на стенках кровеносных сосудов. Определенные изменения происходят также в строении нейрофиламентов центральных нейронов.
Несмотря на огромное число исследований этого заболевания и несомненный прогресс в установлении детальной картины патологических изменений, последовательность развития событий при этом заболевании, точнее, причинно-следственные отношения между установленными симптомами, по-прежнему остаются неясными. Наряду с выраженными явлениями слабоумия и характерной потерей памяти у таких больных находят четкие патологические изменения строения мозговой ткани в ядрах основания переднего мозга — в гигантоклеточном ядре и ядре Мейнерта, откуда идут мощные холинергические проекции в новую кору.
Достаточно убедительно показано, что болезнь Альцгеймера имеет генетическую природу, о чем говорят, в частности, данные семейных исследований, а также молекулярнобиологические исследования последних лет (см. ниже). Белок АРР (amyloid precursor protein) — нормальный компонент нервной системы, синтезирующийся в астроцитах. Ген, кодирующий его, локализован в 21-й хромосоме человека ив 16-й хромосоме мыши. Бета-амилоидный белок, из которого состоят бляшки, характерные для болезни Альцгеймера, есть результат аномального протеолиза АРР-белка.
Экспериментальными генетическими моделями, в которых частично воспроизводится картина нейродегенеративиых изменений, характерных для этого заболевший, являются мыши-трисомики по 16-й хромосоме, а также трансгенные мыши с введенным в геном участком ДНК, кодирующим ген АРР (см,: следующий раздел).
Одной из очень распространенных наследственных болезней человека является так называемый синдром Дауна, как правило связанный с трисомией по 21-й хромосоме, Эта хромосома относится к группе самых мелких аутосом, Синдром Дауна встречается в среднем у одного из нескольких сот новорожденных. Заболевание характеризуется задержкой роста, умственной отсталостью, широким, плоским лицом, "монголоидным" разрезом глаз, рядом аномалий скелета, часто врожденным пороком сердца и предрасположенностью к лейкозам. Мозг больных меньше по размеру, в коре уменьшено число нейронов, а ветвление дендритов более слабое. С возрастом наблюдается ускоренное течение нейродегенеративных процессов в нейромедиаторных системах коры и гиппокампа, в холинергических проекциях ядер основания мозга, в соматостатинергических нейронах коры и в нейронах голубого пятна. У них сокращена продолжительность жизни: чаще умирают в возрасте до трех лет, смертность повышена и в поздние периоды жизни. Считается, что больные с синдромом Дауна быстрее стареют и имеют признаки, характерные для болезни Альцгеймера (см.: 8.6.5.5): амилоидные бляшки и нарушение структуры нейрофибрилл. К 40-летнему возрасту эта патология обнаруживается у 100% больных.
Известно, что с этой трисомией выживает лишь около 30% новорожденных. Вероятность рождения ребенка с синдромом Дауна увеличивается у матерей старше 40 лет (1 новорожденный из 100).
Показано также, что причиной трисомии является нерасхождение хромосом в мейозе, причем относительный "вклад" материнского организма значительно выше, чем отцовского: около 95% случаев связано с дополнительной хромосомой материнского происхождения. Одной из метаболических особенностей болезни Дауна, установленной уже после обнаружения ее генетической природы, является изменение перекисного окисления липидов в клеточных мембранах. В результате этого нарушается процесс инактивации метаболически активных свободных радикалов, высокая концентрация которых служит причиной многих патологических изменений в организме.
Были описаны случаи болезни Дауна, при которых не выявлялась добавочная хромосома. Однако в настоящее время установлено, что каждый больной с этим синдромом имеет, тем не менее, дополнительный генетический материал либо в виде целой 21-й хромосомы, либо в виде ее фрагмента, который называется сегментом 21q22 и может быть перенесен на другую хромосому в результате транслокации.
Хромосома 16 мыши и хромосома 21 человека имеют целый ряд гомологичных элементов, хотя хромосома 16 значительно крупнее, и включает участки гомологии с другими хромосомами человека. Генетическая гомология между этими хромосомами показана, в частности, для участка, где расположена группа генов, в число которых входят гены супероксиддисмутазы и протоонкогена Est-2, a также генов рецептора интерферона клеточной поверхности и фосфорибозилглицеринамидсинтетазы (у человека эти гены картированы в сегменте 21q22, о котором упоминалось выше). Полагают, что сцепление этих генов поддерживается в течение всего времени после расхождения в эволюции "ветвей" человека и грызунов (дата этого расхождения — около 80 млн лет назад).
Специальные экспериментальные приемы позволяют получить мышей-трисомиков по 16-й хромосоме. Этого можно добиться путем использования в скрещиваниях мышей-носителей робертсоновских транслокаций (слияний хромосом), в которые вовлечена 16-я хромосома.
Эмбрионы с трисомией по этой хромосоме нормально появляются на свет, но вскоре погибают. Вес мозга при рождении у них на 20% ниже, чем в норме. Кроме того, для них характерны сходные с синдромом Дауна человека дефекты развития черепа, аномалии сердечной мышцы, иммунной системы, системы кроветворения, а также особенности субклеточного строения нейронов коры. Дифференцировка клеточных элементов мозга у них отстает, а центральные катехоламинергическая и холинергическая системы развиты слабее, чем в норме (Berger et al., 1992).
Поскольку такие мыши-трисомики нежизнеспособны, исследование биохимических путей становления аномального фенотипа на этой модели практически невозможно. Однако оказалось, что можно исследовать мышей-химер (см.: 8.5.3.2), полученных при смешении клеток нормальных эмбрионов и клеток эмбрионов с трисомией 16. Такие животные нормально росли и развивались, нейрогенез у них был описан достаточно подробно. Мыши-химеры, у которых клетки имели либо нормальный кариотип, либо кариотип с дополнительной 16-й хромосомой, отличались от нормальных. У них было меньше нервных клеток, проецирующихся в кору из ядер основания мозга, где основная масса нейронов — холинергические.
Допустимо предположить, что такого рода особенности строения мозга могут оказаться ключевыми в развитии аномалий поведения и при трисомии 16 мыши, и при синдроме Дауна.
По результатам электронномикроскопических исследований корковой пластинки у мышиных эмбрионов с трисомией 16, микротрубочки нейронов аномально закручены, межклеточные контакты недоразвиты, ядра клеток имеют нечеткий контур, а их суммарная площадь уменьшена. Часть этих признаков бывает не только при синдроме Дауна, но и при болезни Альцгеймера.
Как уже говорилось, поскольку существует сильное несоответствие в размерах между хромосомой 21 человека и хромосомой 16 мыши, а также в силу большой отдаленности родства данных видов, эти хромосомы гомологичны друг другу только по небольшому участку. Очевидно, что более адекватной моделью болезни Дауна были бы мыши с тройной "дозой" только данного участка 16-й хромосомы. Такие мыши с трисомией по дистальному участку этой хромосомы получены и исследуются. Показано, что у них повышена экспрессия тех генов, которые у человека "критичны" для развития синдрома Дауна. Эти трисомики обнаруживают и отставание в развитии, и малый рост, и постоянное дрожание всего тела, а также аномалии лицевого черепа. Самцы мышей этого генотипа стерильны. Животные с частичной трисомией находятся в состоянии постоянно повышенного уровня активности и явным образом слабее обучаются навыку отыскания подводной платформы в тесте на пространственную ориентацию (см.: 8.6.7.2).
Перспективной лабораторной моделью для исследования молекулярно-генетических основ синдрома Дауна являются также трансгенные мыши с введенными в геном локусами упомянутого выше участка 21-й хромосомы человека. К концу 1994 г. были получены трансгенные мыши, несущие дополнительные "дозы" гена супероксидцисмутазы (SOD-1), белка S-100B гена APP и ряда других, локализованных в критическом для синдрома Дауна участке 21-й хромосомы человека. У части таких трансгенных мышей были обнаружены четкие симптомы, сходные с теми, которые проявляются при болезни Дауна.
Таким образом, болезнь Дауна как сложнейший симптомокомплекс, затрагивающий формирование многих органов и тканей человека, в том числе и мозга, можно в определенной степени воспроизвести на генетических моделях. Приведенный последним пример "генных" мутаций показывает, что можно добиться даже своеобразного "расчленения" этого синдрома на более простые компоненты.
Это заболевание связано с аномалией X-хромосомы. Оно было обнаружено, когда медики обратили внимание, что в психиатрических больницах число пациентов-мужчин с отставанием умственного развития на 30—50% превышает число женщин с подобной симптоматикой. Анализ родословных выявил, что в этих случаях, как правило, речь идет не о мутации одиночного гена, локализованного на X-хромосоме (наподобие, например, гемофилии), а о принципиально другом явлении. В кариотипе большого числа таких больных-мужчин была обнаружена так называемая "ломкая X-хромосома" (fragile X-chroniosome). На цитологических препаратах это выглядело как сужение или перетяжка на дистальном конце длинного плеча X-хромосомы, причем здесь часто имели место разрывы X-хромосомы. Как только медицинские генетики научились отслеживать этот хромосомный дефект, стало ясно, что данное заболевание распространено очень широко: по частоте встречаемости оно оказалось вторым после трисомии 21 (синдрома Дауна).
В настоящее время неизвестны генетические аналоги этой болезни у животных. Однако нам представляется важным описать ее недавно обнаруженные особенности, поскольку они, по-видимому, достаточно универсальны и могут объяснить сложную картину наследования ряда психических болезней человека. Очевидно, что больной-мужчина получает ломкую X-хромосому от матери. Несмотря на то, что в части тканей матери аномальная по структуре X-хромосома бывает активной (феномен Лайон, см.: Айала, Кайгер, 1988). Однако ни у матерей больных, ни у их родственниц (носителей такой X-хромосомы) заметных отклонений в общем уровне умственного развития поначалу не видели. Однако при более тщательном обследовании большого числа семей с ломкой X-хромосомой были установлены сложные и достаточно уникальные отклонения.
При анализе родословных семей с синдромом ломкой X-хромосомы, у некоторых лиц мужского пола были обнаружены цитогенетические признаки заболевания (в строении X-хромосомы), передавашиеся их дочерям, хотя у них самих признаков психической патологии не было.
Сходные отклонения в "обязательности" передачи по наследству этой патологии свойственны и женщинам. Подобное явление обычно описывается как низкий уровень "пенетрантности" патологического признака (см.: 8.2.6). Забегая вперед, скажем, что в настоящее время оно получает объяснение как феномен предмутационного (потенциального) изменения ДНК *).
*) Премутация, или предмутационное (потенциальное) изменение — генетический феномен, связанный с тем, что лишь небольшая часть повреждений генома превращается затем в мутационные изменения, Большая часть их устраняется в процессе генетической репарации (см.: Фогель, Мотульский, 1990). Наиболее подробно изучены предмутационные изменения при воздействии на геном мутагенных факторов разной природы (Дубинин, 1978; Инге-Вечомов, 1989).
Прогресс в понимании патогенеза данного заболевания стал возможен с применением методов молекулярной генетики (Warren, Ashley, 1995). С помощью генетического и физического картирования, а также генетики соматических клеток в 1991 г. было определено положение данного локуса на X-хромосоме и идентифицирована мутация, ответственная за данный синдром. Ген, получивший название fragile X mental retardation 1 (FMR1), вариабелен по длине: у разных индивидуумов последовательность этого гена включала разное число пар нуклеотидов. Нестабильность размера этого гена определялась вариабельностью длины только одного его участка, который относился не к структурной, а, видимо, к регуляторной части гена. Этот вариабельный участок гена FMR1 состоял из необычно длинной последовательности повторов, состоящих из комбинации (триплета) CGG (цитозин—гуанин—гуанин). Число таких триплетов в обычной популяции человека обычно варьирует от 6 до 54 (в среднем — 30). Далее было показано, что в семьях, где есть больные с синдромом ломкой Х-хромосомы у мужчин-носителей и у их дочерей (у последних не отмечалось присущих этой болезни отклонений 1 психики), число повторов триплета CGG в гене, FMR1 варьирует в пределах 54—200, а у больных их число превышает 200, иногда достигая 1000 и более.
Таким образом, молекулярно-генетическое исследование синдрома ломкой X-хромосомы обнаружило тесную ассоциацию тяжести клинических проявлений болезни и числа повторов триплета CGG. Другое проявление аномалии на молекулярном уровне — это измененный характер метилирования определенного участка данного гена, причем интенсивность нарушения также зависела от тяжести клинической симптоматики. Полагают, что данное нарушение метилирования впрямую связано с изменением (снижением) транскрипции гена FMR1 (Warren, Ashley, 1995).
Итак, существуют какие-то механизмы, посредством которых происходит превращение нормального гена, имеющего малое число повторов специфического триплета, в ген, который еще нормален по своим функциям, но уже легко может стать аномальным. Такая аномалия встречается у лиц-носителей без признаков болезни. Эта измененная последовательность затем преобразуется в такую, которая уже провоцирует функциональные нарушения и может считаться мутантной формой гена.
Итак, исследование синдрома ломкой Х-хромосомы позволило получить конкретную информацию о том, как может происходить превращение премутаций в истинные мутационные события.
Изложенные в предыдущем разделе сведения о молекулярном механизме сложного генетического дефекта — синдрома ломкой Х-хромосомы, были первыми в целой серии экспериментальных открытий молекулярных дефектов при наследственных болезнях человека.
Помимо синдрома ломкой Х-хромосомы такие типы наследственной патологии, как спинальная или бульварная мышечная атрофия, миотоническая дистрофия, болезнь Гентиштона, спинальная и мозжечковая атаксия 1-го типа и денто-рубральная паллидо-льюисова атрофия также связаны с увеличением числа тринуклеотидных повторов. В настоящее время существует мнение о сходном молекулярном механизме проявлений маниакально-депрессивного психоза и шизофрении.
Причины сходства механизмов возникновения патологии при разных заболеваниях нервной системы пока не получили своего объяснения.
Открытие подобных молекулярно-генетических механизмов генеза патологии ЦНС заставляет вспомнить представления отечественного невролога и психиатра С.Н.Давиденкова (см.: 2.5) об особенностях генетически детерминированной патологии человека.
Базируясь исключительно на данных клинических наблюдений, он пришел к выводу, что в широчайшем спектре психических аномалий человека следует различать две принципиально разные категории. Первая из них — это различающиеся по выраженности "вариации нормы", т.е. отклонения, которые могут считаться патологическими, но не обнаруживают признаков глубокого нарушения психики.
Принципиально иной оказывается другая категория — "истинно" патологические состояния, которые характеризуются яркой симптоматикой и не имеют реальной связи с вариациями нормальных черт поведения человека. Тонкий наблюдатель и блестящий клиницист, С.Н.Давиденков смог обратить внимание на принципиальное различие патологий этих двух типов. Нетрудно видеть, что его подразделение аномалий психики человека оказывается аналогичным описанным выше явлениям — существованию "уже аномальных" генотипов, в которых мутация не получила полного развития, и действительно мутантных генотипов, в которых дефект можно зафиксировать и на уровне фенотипа, и на молекулярногенетическом уровне.
Как уже упоминалось, огромное большинство признаков поведения характеризуется непрерывной изменчивостью, и подобные ряды нельзя подразделить на фенотипически дискретные классы. Такой характер фенотипической изменчивости можно объяснить одновременным действием большого числа генов в сочетании с фенотипическими отклонениями, которые вызываются влиянием средовых факторов.
В разделе, посвященном проблемам нейрогенеза (см.: 8.5.1), говорилось о компонентном составе фенотипической изменчивости в применении к проблеме относительной роли генотипа и среды при формировании признака. При оценке количественных признаков сформированного организма выступают сходные по структуре, но несколько иные по природе закономерности. Ниже будет дан их анализ.
Вклад генетической изменчивости в фенотипическую подразделяется на три части: аддитивные генетические эффекты, отклонения доминирования и эпистатические взаимодействия. Основная модель количественной генетики представлена на рис. 8.24.
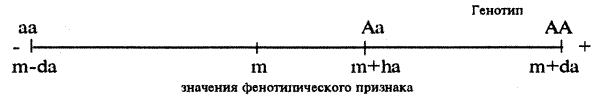
Рис. 8.24. Схематическое изображение возможных эффектов действия одиночного гена, имеющего два аллеля. Параметр da — величина, равная половине разницы между двумя гомозиготами; ha — величина, на которую признак отклоняется у гетерозиготного генотипа от точки m, — это средняя величин признака обоих родителей. В генетике количественных признаков заглавными буквами обозначают усиливающие аллели, которые не обязательно доминантны. Таким образом, величина da положительна по определению, тогда как ha может принимать любые значения
Если мы возьмем инбредную линию, гомозиготную практически по всем локусам, поскольку она прошла много поколений братско-сестринских скрещиваний, то фенотипический признак, определяющийся большим числом генов, можно обозначить как m + (d+) + (d-) ; где (d+) — суммарный эффект генов, представленных в данном фенотипе усиливающими аллелями; (d-) — то же для ослабляющих аллелей; параметр m — постоянная, которая отражает средние средовые эффекты, общие у двух сравниваемых линий, а также генетические эффекты для локусов, в которых сравниваемые линии имеют одинаковые аллели.
Различия в величине признака для двух инбредных линий будут выражаться как 2[(d+) + (d-)]; эту величину мы будем обозначать для краткости как 2[d]. Точно также величина признака у гибрида первого поколения выразится как m + (h+) + (h-), далее обозначается как m + [h].
Отметим, что А — это суммарное, связанное с доминированием, отклонение для многих генов. Если эффекты этих генов противоположны по направлению, то [h] будет малым или равным нулю, даже если доминирование имеет место. То же относится и к [d]: даже когда у двух инбредных линий, несущих разные аллели в существенных для признака локусах, фенотип определяется аддитивными генетическими эффектами, межлинейные различия необязательно будут заметными. С другой стороны, у гибридов может возникнуть ситуация, когда две родительские линии имеют доминантные аллели, равно распределенные между ними. Тогда в случае направленного доминирования (т.е. когда все отклонения доминирования имеют одно направление) все гибриды будут обладать крайними значениями признака. Из-за этого фактическая величина [h] может оказаться больше, чем [d]. Это явление называется гетерозис.
Сложнее описать участие генотипа в определении изменчивости признака при эпистатических (т.е. межлокусных, или неаллельных) взаимодействиях. Этими терминами обозначают явления, когда эффекта разных генов не суммируются простым образом, а фенотипический эффект аллелей одного локуса зависит от генотипа другого локуса. Наиболее простой и известный пример эпистатического взаимодействия — это взаимодействие локусов, влияющих на окраску шерсти у мышей. Например, локус b определяет черный или коричневый цвет шерсти. Однако, если мыши гомозиготны по локусу альбинизма с, то пигмент просто не образуется и никакие аллельные замены в локусе b ничего изменить не смогут, Эпистатические взаимодействия элементов полигенной системы настолько сложны, что ее точное описание невозможно, Для простоты описания обычно постулируется отсутствие значимых межаллельных взаимодействий. Более подробное знакомство с методами генетики количественных признаков помогает в оценке результатов генетических исследований признаков поведения и позволяет построить модели, которые описывают достаточно сложные генетические феномеиы, такие как, например, плейотропия.
Важным понятием в генетике количественных признаков является также наследуемость "в узком" и "в широком" смысле. Наследуемость в узком смысле определяют как долю всей фенотипической дисперсии, которая определяется генами с аддитивными эффектами. Наследуемость в широком смысле — это доля всей генетической изменчивости в фенотипической.
Из этих определений следует, что величина наследуемости зависит от генетических особенностей особей изучаемой популяции, от средовых условий, а также от степени точности, с которой определяется фенотип.
Можно кратко проанализировать случай, когда необходимо оценить корреляцию между двумя признаками, каждый из которых находится частично под контролем общих генов. Иными словами, в данном случае нас больше интересует ковариация между признаками, чем просто их изменчивость. Фенотипическая коварианса двух признаков имеет генетическую и средовую составляющие. В свою очередь генетическая составляющая может быть подразделена на компоненты, связанные с аддитивным эффектом и с эффектом доминирования.
Если мы определили величины генетических коварианс и дисперсий (варианс), то вслед за этим можно вычислить генетическую корреляцию между признаками. Отметим, что подобные расчеты проводятся на основе математических моделей со значительным числом допущений (например, об отсутствии неравновесия при сцеплении см.: Айала, 1988). Если фенотипические корреляции между признаками могут оказаться следствием целого ряда случайностей и/или побочных эффектов, то генетические корреляции отражают реальное существование "общего пути" в формировании двух признаков, иными словами, реальное существование общих генов, которые детерминируют интересующие нас признаки.
Фенотипическая изменчивость есть не только сумма эффектов изменчивости генотипа и среды, она включает в себя также величину, связанную со взаимодействием этих факторов. На основе этого соотношения были разработаны методы оценки наследуемости, степени доминирования аллелей, а также так называемые многопараметрические статистические методы в применении к генетике количественных признаков (см.: Мазер, Джинкс, 1985; Crusio, 1992).
Для популяционной генетики, этологии и общей биологии генетика количественных признаков важна как способ оценки так называемой "генетической архитектуры признака". При анализе поведения животных исследователь, как говорилось выше, руководствуется одним из четырех основных вопросов, которые в наиболее четкой форме были сформулированы Тинбергеном (Tinbergen, 1963).
Два из них относятся к "непосредственным причинам" наличия данного признака в репертуаре поведения, т.е. к его нейрофизиологическим механизмам и/или его развитию в онтогенезе, два других касаются: один — функциональной значимости признака в настоящий момент, т.е. его адаптивной роли, а другой — его роли в процессе эволюции вида, т.е. в филогенетическом аспекте. Два вопроса "функциональной" группы часто называют анализом причин (causation) данного поведения в филогенетическом и феногенетическом аспектах. Генетическую архитектуру признака поведения также можно рассмотреть с этих позиций.
Филогенетический аспект. Как правило, для подобного анализа необходима информация о составе генетической изменчивости. В соответствии с генетической теорией естественного отбора (Fisher, 1958; Айала, 1988) отбор на высокие величины какого-либо признака имеет результатом низкий уровень его аддитивной генетической изменчивости и большую величину генетической изменчивости, связанной с доминированием. Для подобных признаков величина наследуемости в узком смысле бывает невысокой, в отличие от значительной наследуемости (в широком смысле).
В случае стабилизирующего отбора, т.е. отбора на средние значения признака, его генетическая архитектура будет характеризоваться большой величиной аддитивного компонента изменчивости (дисперсии) и либо отсутствием доминирования, либо двусторонним доминированием.
Наследуемость в узком смысле будет в таких случаях высокой, а доминирование — относительно малым.
Оценка признака поведения в условиях лабораторных тестов зависит, как правило, от многих переменных (не имеющих, разумеется, экологической ценности). Они могут дать некую величину аддитивной изменчивости, что, однако, ни в какой мере не может быть индикатором какой бы то ни было адаптивной значимости признака (как это принято считать для "естественных" нелабораторных признаков).
В этом отношении интересны данные американского исследователя Н.Хендерсона (Henderson, 1978). Уровень двигательной активности 12-дневных мышат разных генотипов он оценивал в двух ситуациях: при помещении мышонка в установку "открытое поле", т.е. в типичном лабораторном тесте, и при "возвращении" вынутого из гнезда детеныша назад, в привычные комфортные условия. Оказалось, что аддитивная компонента дисперсии признака "уровень двигательной активности", т.е. быстроты перемещения животного, была высокой в первом, и низкой — во втором случае. Двигательная активность животного в разных условиях контролируется разными физиологическими механизмами. Возвращение мышонка в гнездо — реакция с четкой адаптивной функцией. Низкие величины аддитивного компонента дисперсии этого признака свидетельствуют о том, что формирование его в ходе эволюции происходило с отбором на крайние значения (большую скорость перемещения). В то же время "активность в открытом поле" (т.е. выполнение практически тех же движений, но, видимо, с иным мотивационным фоном) имеет другую генетическую архитектуру с большой величиной аддитивной компоненты дисперсии. Возможно, что последние результаты связаны с неадекватным характером теста "открытое поле" для детенышей столь раннего возраста.
В этом контексте следует упомянуть еще об одном понятии, которое часто используется при обсуждении адаптивной и/или функциональной роли признака — о так называемой приспособленности организма, т.е. о его способности выжить и оставить способное к размножению потомство.
Признаки, имеющие высокую величину компоненты доминирования в величине общей генетической дисперсии, связаны с приспособленностью.
Естественный отбор благоприятствовал сохранению особей с наибольшими величинами такого признака. В то же время затруднительно делать подобные выводы о признаках с двусторонним доминированием, т.е. о тех, у которых предположительно был отбор на средние значения. В таких случаях наиболее благоприятными для выживания могли оказаться средние значения признака. Очевидно, что разделение признаков, влияющих и не влияющих на приспособленность, достаточно условно, поскольку базируется на результатах вычислений с целым рядом допущений.
Феногенетический аспект. Этот подход мы обозначили выше как путь "от гена к признаку". В его рамках проводятся исследования корреляций между поведением и физиологическими, нейроанатомическими и нейрохимическими переменными, которые могут пролить свет на происхождение этих связей (см.: 8.6.7.2).
Метод выявления генетических корреляций позволяет обнаружить совместное варьирование генетических эффектов. Такие корреляции выражают либо плейотропные эффекты (см.: 8.2.2), либо так называемое неравновесие, или неравновесность, по сцеплению*) (linkage disequilibria; см.: Айала, Кайгер. 1988). Если для анализа генетических корреляций выбирать неродственные между собой группы, например инбредные линии разного происхождения, то вероятность неравновесности по сцеплению будет невелика, поскольку, если у них и существуют "коадаптированные локусы", у гибридов они не сохранятся. В этом случае генетическая корреляция между двумя признаками будет свидетельствовать о том, что, по меньшей мере, часть пути "от гена к признаку" для этих двух признаков оказывается общей.
*) Термином "неравновесие по сцеплению" называют явление, когда какое-то сочетание аллелей двух разных локусов дает более "приспособленный" фенотип, чем сочетание других аллелей этих же локусов. На практике это может означать неслучайную ассоциацию признаков в данном генотипе, причем это не связано с общими физиологическими механизмами их реализации.
Это делает достаточно высокой вероятность существования между ними функциональной связи. Напомним, что наличие фенотипической корреляции между признаками еще не может быть указанием на общность их генетической детерминации (см.: 8.6.1). Корреляционные решетки, полученные для величин генетических корреляций, можно далее анализировать с использованием стандартных многопараметрических методов, например факторного анализа.
Конкретные методы оценки состава генетической изменчивости и величин наследуемости достаточно многочисленны, и рассмотреть их здесь не представляется возможным (см.: Эрман и Парсонс, 1984; Мазер, Джинкс, 1985). Наряду с определением регрессии родители—потомки, а также селекции линий на разные величины признака, в генетике поведения большое распространение получили методы анализа инбредных линий. Один из таких методов — так называемое диаллельное скрещивание.
Это один из наиболее употребительных приемов генетического анализа при исследовании генетической детерминации признаков поведения. Суть его заключается в оценке средних величин признака и их дисперсии у животных нескольких (минимум трех) инбредных линий, а также гибридов всех возможных сочетаний. В результате такого скрещивания получается p2 групп, различающихся по генотипам, где р — число исходных линий. На животных в соответствующем возрасте измеряют интересующие нас признаки. Очевидно, что условия содержания таких животных должны быть идентичными. Кроме того, непременное условие диаллельных экспериментов — это применение методов рандомизированных блоков: каждый вариант скрещивания должен быть осуществлен как минимум дважды.
Диаллельный анализ — это вариант дисперсионного анализа, правила которого изложены в специальных руководствах (см.: Heyman, 1954; Турбин и др,, 1974), В зависимости от задачи, стоящей перед исследователями, для диаллельного анализа можно применять разные теоретические модели. В любом случае результатом проведения подобного исследования бывает получение величин компонент дисперсии, а также величин, характеризующих уровни ковариации признаков у разных генетических групп, участвующих в эксперименте.
Первое исследование такого рода провели при скрещивании шести инбредных линий крыс, проанализировав уровни дефекации и двигательной активности в "открытом поле". Для уровня дефекации наследуемость в узком смысле (см.: 8.6.6.1) оказалась равной 0,62, а для уровня двигательной активности (оценивалась по числу пересеченных квадратов) — 0,59. Это означает, что в условиях лабораторного тестирования в установке "открытое поле" аддитивная компонента дисперсии для этих признаков достаточно велика (Бродхерст, 1975).
Ряд примеров использования данного метода в работах по генетике поведения дрозофилы можно найти в книге Эрман и Парсонса (1984).
Метод диаллельного скрещивания имеет ряд преимуществ по сравнению с классическим генетическим анализом. Первое из них состоит в том, что проведение такого эксперимента занимает меньше времени, поскольку можно ограничиться исследованием признака у первого поколения гибридов. Группа (популяция) исследуемых животных позволяет измерять нужный признак и у родительских генотипов — инбредных линий, и у их потомков-гибридов, что бывает очень важно при исследовании признаков поведения, которые зависят от сезона, температуры, особенностей питания и других факторов. Третье преимущество заключается в том, что потомки диаллельного скрещивания представляют собой популяцию с известным генетическим составом, которая может служить для оценки генетических (и микроэволюционных) эффектов тех или иных экологических факторов. П.М. Бородин (1981) дал краткое описание статистических методов анализа диаллельных экспериментов, а также интерпретировал его результаты.
Помимо данных о характере генетической изменчивости признака поведения, результаты диаллельного эксперимента иногда дают информацию, существенную для понимания физиологических процессов, лежащих в основе этого признака. Обратимся вновь к психофизиологическим особенностям условной реакции избегания у крыс и мьшей в челночной камере, когда животные вынуждены возвращаться в отсек, где они только что получили болевое раздражение (см.: 8,6.3.2), По данным диаллельного скрещивания, проведенного Дж, Вилкоком и Д, Фалкером, т.е. при сопоставлении генетической изменчивости показателей обучения у родительских линий и соответствующих гибридных животных, картина оказалась сложной, Низкий уровень выполнения реакции избегания на первых этапах эксперимента имел доминантное наследование (см,: Вилкок, 1975), Иными словами, аллели, определяющие замедленную двигательную реакцию животного в ответ на боль в первые дни обучения, были доминантными. На более поздних этапах выработки условной реакции доминантное наследование обнаруживается у аллелей, определяющих высокую частоту выполнения навыка. В то же время следует отметить, что у животных некоторых исследованных генотипов имелись аллели, определявшие быструю реакцию уже в первые дни обучения.
Итак, казалось бы, внешне (фенотипически) один и тот же признак поведения — выполнение реакции избегания в челночной камере — на разных стадиях обучения находится под генетическим контролем разных систем, или, другими словами, на разных этапах выработки этой реакции задействованы разные физиологические процессы. Именно этот вывод и сделали авторы, предположив существование двух физиологических систем, влияющих на признак в начальной и конечной стадии формирования реакции избегания, генетический контроль которых осуществляется независимо.
Авторы этого исследования полагали, что генетическое разнообразие в частоте выполнения реакции на первой стадии обучения, по-видимому, связано с изменчивостью порогов реакций животных на болевую стимуляцию (затаивания или бегства), где доминантной была тенденция к затаиванию. По мере формирования ассоциативной связи в ходе повторения сочетаний условного и безусловного раздражителей ситуация меняется, и генетическое разнообразие по измеряемому признаку теперь уже связано с изменчивостью в скорости формирования условной связи и/или в выполнении навыка (т.е., видимо, с изменчивостью собственно способности к обучению).
Исследование генетических закономерностей наследования количественных признаков в селекционных экспериментах и при сравнении инбредных линий обеспечивает как бы базис для оценки наследования сложных признаков поведения. Эти методы не могут ответить на вопрос о числе генов, обеспечивающих генетическую изменчивость по данному признаку и, тем более, об их локализации.
Метод рекомбинантных инбредных линий (РИЛ), разработанный Н. Бейли еще в 70-е годы (Baily, 1971), в целом ряде случаев позволяет выявить участие небольшого числа "главных" генов (т.е. генов, которые обеспечивают наибольший вклад в изменчивость данного полигенного признака), а также дать информацию об их локализации на хромосоме.
Рекомбинантные инбредные линии не следует путать с линиями коизогенными (идентичными по генотипу, за исключением некоего локуса, в котором произошла мутация) и линиями конгенными (идентичными по генотипу за исключением некоего локуса с геном-маркером, введенным в генотип специальными скрещиваниями), которые были кратко описаны выше (см.: 8.6.4.1).
РИЛ получают на основе популяции гибридов второго поколения, в свою очередь полученных от скрещивания гибридов первого поколения двух инбредных линий, контрастных по изучаемому признаку. Инбредные линии, как известно, гомозиготны по всем локусам. Гибриды первого поколения двух неродственных между собой инбредных линий гетерозиготны по всем аллелям, а все особи генетически идентичны друг другу. У гибридов второго поколения происходит расщепление (1:2:1) по каждому гену. Путем инбридинга (братско-сестринского скрещивания), начатого с особей-гибридов второго поколения и осуществляющегося в течение обычно 10—12 поколений, получают серию новых инбредных линий, гомозиготных по всем локусам. Это и есть рекомбинантные инбредные линии. Каждая из них представляет собой "зафиксированный" в гомозиготном состоянии один из вариантов расщепления признаков, т.е. результат серии событий расщепления и рекомбинации при формировании гибридов второго поколения.
Если про обычные инбредные линии можно сказать, что каждая из них представляет собой некий гомозиготный генотип, повторенный неограниченное число раз, то каждая из РИЛ данной серии — это также воспроизведенный многократно и зафиксированный в гомозиготной форме вариант перекомбинации аллелей двух линий в том виде, как это произошло у гибридов второго поколения. При этом, поскольку внутри каждой линии все животные идентичны друг другу генетически, линии можно сравнивать как между собой, так и с родительскими линиями.
В случаях, когда взятые в исследование исходные инбредные линии различаются по аллелям какого-то интересующего нас гена, половина новых линий будет гомозиготна по одному его аллелю (от родителя А), а вторая половина — по другому аллелю (от родителя В). Ясно, что в этом случае мы можем обнаружить две группы (два "кластера") новых линий. Если эти кластеры по значению интересующего нас признака не перекрываются между собой, это четко свидетельствует о его моногенном наследовании, а именно такие случаи можно анализировать далее методом РИЛ. И наоборот, если у набора РИЛ обнаруживается непрерывное распределение значений признака, это свидетельствует о полигенном характере наследования. В последнее время разработан метод, позволяющий анализировать и такие случаи. Это метод картирования QTL (см.: 8.6.6.4).
В случаях, когда данная группа РИЛ, тестированная на проявление интересующего нас признака, показывает бимодальное распределение его величин, т.е. если признак имеет моногенное наследование, вступает в действие следующая процедура. Характер распределения значений изучаемого признака в нашей группе РИЛ (в англоязычной литературе это — strain distribution pattern, SDP), сопоставляется с SDP других признаков, которые контролируются уже ранее картированными генами, т.е. генами, чья локализация на хромосомах известна (часто это гены тканевой совместимости), и которые в данном случае выполняют роль маркеров.
Если такие SDP совпадают, имеются основания говорить о локализации гена, ответственного за формирование интересующего нас признака достаточно близко к маркеру. Близкое расположение этих двух генов предполагается потому, что в ходе гибридизации и последующего инбридинга хромосомы претерпевают несколько кроссинговеров, и сходное распределение двух генов — сходные SDP- могут обнаружиться только при близком их топографическом расположении. В пионерской работе Д. Бейли (Bailey, 1971) описывался метод создания РИЛ на основе линий BALB/c и C57BL/6 (серия СхB). В дальнейшем были получены и другие серии линий. Это в первую очередь выведенная Б. Тэйлором серия из 26 РИЛ на основе линий C57BL/6 и DBA/2 (серия BxD), а также линии, которые были выведены американской исследовательницей М. Несбит на основе реципрокных гибридов между C57BL/6 и А (Ас4 и АхB) (Nesbit, 1992).
Очевидно, чем большее число линий содержит серия, тем более информативными оказываются результаты исследования. Бейли удалось вывести всего 7 РИЛ, тогда как в серии BD их 26. Эти РИЛ успешно используются в работах по локализации генов, связанных с формированием алкоголизма у мышей. На апрель 1992 г. для этой серии РИЛ было выявлено 550 маркерных SDP, в том числе 240 SDP, которые уникальны именно для этой серии. Общая длина ге-нома мыши составляет 1600 сантиморганид (морганида — единица генетического расстояния, измеряемая процентом кроссинговера, 1 морганида — отрезок хромосомы, в пределах которого наблюдается 1% перекреста). При наличии 240 SDP уникальных для этой комбинации линий в среднем на каждые 6,6 сМ имеется четкий маркер. Это означает, что при использовании РИЛ серии BxD каждый из описанных новых локусов с большой вероятностью может быть кар-тирован. Мощность метода РИЛ усиливается применением молеку-лярнобиологических методов, позволяющих выявлять множество новых маркеров, например минисателлитные и микросателлитные повторы, а также полиморфизм длины рестрикционных фрагментов (Льюин, 1987).
В последние годы, в связи с интенсивным исследованием генома мыши методами молекулярной биологии (в частности, с использованием техники PLRF — полиморфизма длины рестрикционных фрагментов) была накоплена обширная библиотека данных по молекулярным маркерам у данного вида животных. Это означало, что практически по всей длине ДНК мыши с достаточно малыми интервалами имелись специфические последовательности оснований, наличие или отсутствие которых можно было определить экспериментально. В частности было установлено, что ряд линий мышей имеют достоверные и четкие различия по подобным молекулярным маркерам, что позволило разработать метод картирования локусов количественных признаков — QTL
Впервые картирование "локусов количественных признаков" т.е. локусов, каждый из которых вносит свой небольшой вклад в признак, доступный количественной оценке, впервые осуществили Дж. Маклерн и Р. Пломин с сотр. в начале 90-х годов с использованием батареи РИЛ. Кратко изложим суть подхода. Выбранный молекулярный маркер (т.е. определенная последовательность оснований, которую можно увидеть при электрофорезе ДНК животного данного 346 генотипа), присутствует у одной из родительских линий, основопо-ложниц данной группы РИЛ, и отсутствует у другой. Как и у F2 от скрещивания этих линий, он обнаруживается и у некоторых линий данной группы РИЛ. Далее у всех линий данной группы РИЛ можно вычислить корреляцию наличия данного маркера со средними величинами исследуемого количественного признака. Вычисленный таким образом простой коэффициент корреляции будет равен нулю для аллелей одной из родительских линий, и единице — для другой. Величина коэффициента показывает значимость различий между средними величинами признака для двух генотипов, а также указывает на силу связи. Умеренная величина корреляции может быть результатом либо тесной связи маркера с локусами, незначительно влияющими на данный признак, либо слабой связи с подобными локусами, которые оказывают большой эффект. Какая из этих альтернатив верна, можно определить при дальнейшем анализе. При анализе данной группы "количественных локусов" коэффициенты корреляции вычисляются для каждого из нескольких маркеров, расположенных на некотором участке хромосомы. После этого можно обнаружить, что величина коэффициентов для маркеров вдоль всего хромосомного фрагмента меняется. Из этого можно сделать вывод о том, где пространственно локализуется один из локусов, вносящий свой вклад в исследуемый количественный признак.
Напомним, что метод РИЛ оказывается достаточно эффективным, если стоит задача локализации гена, контролирующего данный признак "в одиночку", или если на признак преимущественно (это так называемые главные гены) влияет именно этот ген. В таких случаях рекомбинантные линии различаются между собой по исследуемому признаку. Однако метод QTL, основанный на использовании огромного количества маркеров, обладает еще большей разрешающей способностью по сравнению с РИЛ в его изначальном варианте. Картирование локусов количественных признаков позволяет исследовать предполагаемые генетические различия по некоторому признаку даже в случае, если между линиями нет фенотипических различий.
Поясним сказанное. Величины количественного признака у двух линий могут быть одинаковыми, но наборы генов с малыми эффектами, из которых эти величины складываются, не обязательно идентичны. Метод QTL обладает достаточной "мощностью" для выявления таких различий. Очевидно, что физиологические признаки, которые подвержены множественным регуляторным воздействиям и в зависимости от генотипа в разной степени подвержены внешним влияниям, могут быть с успехом проанализированы методом QTL Примером может служить обнаружение локусов, участвующих в обеспечении реакции животного на помещение в установку "открытого поля" (Genshenfeld et al., 1997).
Внутривидовая изменчивость строения мозга затрагивает его абсолютный размер, относительную величину, а также количественную изменчивость отдельных мозговых образований. Показано, что у мышей гетерогенных популяций и у представителей инбредных линий абсолютные и относительные размеры мозга сильно варьируют. Так, например, у линии DBA/2J средняя масса мозга достаточно мала и сильно отличается от средней величины, полученной для всех исследованных линий, а у линии BALB/c она наибольшая: соответственно 430 и 550 мг.
Оценки внутрилинейной и межлинейной изменчивости в ряду 25 инбредных линий позволили определить, что вариация веса мозга у мышей имеет генетическую компоненту. Это было подтверждено и в нескольких успешных экспериментах по селекции мышей на большую и малую массу мозга (Wimer et al., 1969; Попова и др., 1976; 1996; Fuller, 1989). Остановимся несколько подробнее на одном из таких экспериментов, проведенном в лаборатории физиологии и генетики биологического факультета МГУ (Попова и др., 1977). Генетически гетерогенная популяция, на основе которой был начат селекционный эксперимент, была получена в результате скрещиваний в течение 4 поколений мышей нескольких инбредных линий ( СВА, DBA/2J, C57BL/6J, C57BR/cd, BALB/c к А/Не).
Отбор животных для получения каждого последующего поколения проводили на основе линии регрессии, построенной для двух скоррелированных признаков — веса тела и веса мозга у взрослых особей. Половину особей каждого помета забивали и определяли у них вес мозга и вес тела. Если большая часть точек (каждая из которых отражала данные по одному животному) попадала за пределы доверительного интервала при данном весе мозга, то остальных мышат этого помета брали на скрещивание для получения следующего поколения линий, соответственно с большим или малым весом мозга. В тех случаях, когда у мышей одного помета значения веса мозга группировались вблизи линии регрессии или имели значительный разброс, весь помет из дальнейшей селекции исключался. Во всех последующих поколениях селекции линию регрессии рассчитывали отдельно для каждой линии.
Межлинейные различия в весе мозга во всех селекционных экспериментах были достоверны, их абсолютная величина составляла 70-А 90 мг, т.е. примерно 15—20% от среднего веса мозга. Отметим, что при подобной селекции отмечались также и скоррелированные изменения в весе тела, но, поскольку критерии отбора были основаны не на абсолютных величинах, а на положении соответствующих точек на линии регрессии, различия оставались незначительными и сопоставимыми по величине с естественными вариациями признака.
Какие же изменения в строении и функции мозга вызывает подобная селекция? На основании нескольких селекционных экспериментов можно считать установленным, что мыши, селектированные на большой вес мозга, обладают более высокой способностью к обучению. Эти различия были обнаружены при обучении животных на основе пищевой мотивации, а также при выработке реакции избегания удара электрического тока.
Более высокую обучаемость мышей с большим весом мозга, таким образом, нельзя объяснить случайной ассоциацией генетических факторов, возникшей в ходе искусственного отбора, поскольку данный эффект наблюдался в разных парах селектированных линий. Темпы созревания рефлексов в раннем онтогенезе также имели устойчивые различия: мышата из линий с малым весом мозга развивались быстрее (Попова, 1986). Возможно, что более высокая способность к обучению как проявление большей пластичности ЦНС может оказаться скоррелированной с более медленными темпами созревания нервных клеток и межклеточных контактов.
Иными словами, оба эти свойства мозга могут находиться под влиянием каких-то сходных факторов, выраженность которых у мышей, селектированных на большой вес мозга, оказывается устойчиво более высокой. Еще одним устойчивым различием, проявившимся по крайней мере в двух селекционных экспериментах, была более высокая судорожная готовность мышей, селектированных на малый вес мозга. Как при судорогах, вызываемых ударом тока, так и при судорогах, спровоцированных фармакологическими агентами — пентилентетразолом и стрихнином, мыши с малым весом мозга были достоверно более чувствительными (см.: Богданов и др. 1994). Причины этого феномена остаются неясными, а их исследование может быть важным для выявления генетического контроля эпилептогснеза в целом (см.: 8.6.5.1).
Морфологические и цитоархитектонические исследования различий между данными линиями в эксперименте Н.В. Поповой выявили одно чрезвычайно интересное обстоятельство. У мышей, селектированных на большой вес мозга, площади новой, старой и древней коры были больше, а новая кора отличалась особым типом "упаковки" клеточных элементов. У мышей с большим весом мозга корковые клетки располагаются в виде более плотных скоплений, разделенных пространствами с реже расположенными клетками. Возможно, что эта особенность строения мозга мышей, селектированных на его большой вес, каким-то образом связана с их более низкой подверженностью судорогам, о которой говорилось выше (Попова и др., 1983).
В 1971 г. была опубликована работа американских исследователей Р. и Ц. Ваймеров и Т. Родерика (Wimer et al., 1971), в которой они сопоставили способность мышей генетически гетерогенной популяции к обучению пассивной реакции избегания удара электрического тока (при единственном предъявлении) с размерами гиппокампа. Далее мозг этих животных исследовали методами морфометрии для определения объема гиппокампа. Оказалось, что чем больше был размер гиппокампа, тем эффективнее было пассивное избегание у данного животного, Таким образом, впервые была установлена достоверная положительная корреляция между изменчивостью этой структуры мозга и изменчивостью поведения.
Исследования в этом направлении были продолжены, Сопоставление нейрогистологических препаратов мозга большого числа линий и популяций мышей и крыс выявило межлинейные различия целого ряда отделов гиппокампа. Встал вопрос о реальном существовании связи между строением гипппокампа и поведением. Эти исследования были начаты в 70-х годах Х.-П, Липпом и соавт, (см.: Lipp etal,, 1989), Первоначально для такого анализа была выбрана наиболее популярная в то время условная реакция избегания в челночной камере. Выше (см,: 8,6,3.2) упоминалось о недостатках данной методики.
Индивидуальную изменчивость темпов обучения этой реакции исследователи рассматривали как зависимую переменную, тогда как в качестве независимой переменной взяли вариабельность площади терминального поля — зоны окончания мшистых волокон, аксонов гранулярных клеток зубчатой фасции гиппокампа на апикальных дендритах пирамидных нейронов поля САЗ. Мшистые волокна оканчиваются в пирамидном слое крупными синаптическими бляшками. Зоны окончаний формируют два четких синаптических поля, две области проекции. Одна из них располагается непосредственно над пирамидными нейронами поля САЗ и называется супрамирамидным слоем. Вторая, меньшая по объему, располагается ниже или внутри слоя пирамидных клеток. Эта область называется слоем интра- и инфрапирамидных мшистых волокон, iipMF. В дальнейшем были разработаны методики определения размеров этой области. На основе количественных данных по площадям данной области на серийных горизонтальных срезах мозга, окрашенных по методу Тимма (выявление цинксодержащих участков нейронов гиппокампа), можно определить объем всей структуры (Lipp, Wolfer, 1995).
В исследованиях X.-П. Липпа и его коллег быстроту формирования навыка избегания удара электрического тока в челночной камере сопоставляли с размером именно этой изменчивой структуры мозга у животных разных генетических групп. Объектами с высоким уровнем генетической и фенотипической изменчивости размеров iipMF были селектированные линии крыс с четкими различиями поведения и инбредные линии мышей с разной скоростью обучения навыку избегания тока. Было проанализировано также и поведение гибридных животных, полученных от скрещивания линий, контрастных по изучаемым признакам. Кратко дадим результаты этой серии исследований (Lipp et al., 1989).
Селектированные линии крыс. Размер iipMF оценивали у двух линий крыс, селектированных на высокую и низкую способность к обучению избеганию в челночной камере — линии Roman High (RHA) и Roman Low Avoidance (RLA) (см.: 8.6.3.2).
Строение гиппокампа у этих линий различно. У RHA были толще поверхностный слой moleculare-lacunosum и слой базальных дендритов stratum oriens. У RLA была больше площадь обеих зон окончаний мшистых волокон. Иными словами, обнаруживалась отрицательная корреляция между площадью проекции мшистых волокон и способностью к обучению реакции избегания тока в челночной камере.
Таким образом, в результате селекции крыс по способности к обучению данной реакции в этих линиях произошли изменения морфологии гиппокампа. Следует помнить, что подобная ассоциация признаков, возникшая в результате селекции, может быть случайной, т.е. эти признаки могут быть и не связанными друг с другом общей причиной. Выше (см.: 8.6.2) упоминалось, что, поскольку при селекции обычно используется малое число животных, может произойти случайное закрепление в линии того или иного сочетания несцепленных между собой аллелей. Чтобы выяснить, не является ли подобная корреляция случайной, необходимо либо повторить селекцию, либо провести дополнительный анализ связи обнаруженных феноменов на других объектах, либо проанализировать корреляцию признаков у гибридов ''расщепляющихся поколений'' (F2, РЗ и др.).
Инбредные линии мышей. У 7 инбредных линий — DBA/2, СЗН/Не, BALB/c, C57BL/C, NMRJ, ICR, SM/J - также проанализировали корреляцию площади терминальных полей мшистых волокон и показателей поведения в челночной камере. Сильная и высокодостоверная отрицательная корреляция была выявлена между размером iipMF и обучаемостью: -0,92. Корреляций с размерами других структур гиппокампа не обнаружилось.
Рандомбредная популяция мышей. Итак, если такая корреляция действительно отражает причинную связь между размером данного синаптического поля и обучаемостью в челночной камере, то подобную связь можно найти и в генетически гетерогенной популяции животных, полученной в результате случайного скрещивания нескольких контрастных по этим признакам генотипов. Эксперименты это подтвердили: величина корреляции была -0,82.
Уже совокупность приведенных фактов позволила предположить существование причинной связи между особенностями строения гиппокампа и поведением, однако были получены и другие подтверждения такой ассоциации признаков.
Линии крыс В, Н, ВЛ н НЛ. В Институте физиологии им. Павлова АН СССР к середине 70-х годов существовали две пары селектированных линий крыс. Первая пара линий ( В и Н) была выведена с отбором на высокий и низкий пороги нервно-мышечной возбудимости, вторая (ЕЛ, ЯД или КНА и KLA, см.: 8.6.3.2) — на скорость образования условной реакции активного избегания в челночной камере. Оказалось, что у крыс линии НЛ площадь iipMF была выше, чем у контрастной линии. В то же время крысы, селектированные на разную величину порога нервно-мышечной возбудимости (В и Я), не различались по площади проекции, но обучались с разной скоростью: у крыс Н обучение шло успешнее. Этот результат, разумеется, не был неожиданным, поскольку такой сложный признак, как обучение избеганию удара тока может зависеть и от характеристик периферической нервной системы.
Крысы, селектированные на разную частоту самостимуляции. Исследование 2 пар таких линий показало, что они различаются по размеру iipMF и по уровню реакции избегания, и эти признаки также оказались скореллированными.
Мыши инбредных линий. Генетический анализ. Ранее было показано, что инбредные линии мышей DBA/2 и СЗН контрастны и по величине iipMF, и по способности к обучению избеганию. В специальном эксперименте были получены гибриды второго поколения от скрещивания этих линий и проанализированы индивидуальные корреляции все тех же двух признаков. Высокая отрицательная корреляция размера площади проекции iipMF и успеха обучения в челночной камере была обнаружена и в этом исследовании.
Мыши инбредных линий. Диаллельное скрещивание. Как рассматривалось выше (см.: 8.6.6.2), в таком эксперименте анализируют признаки у гибридов первого поколения от скрещивания животных нескольких генотипов. При этом изменчивость признака внутри группы (т.е. между особями с идентичным генотипом) связана с влиянием средовых факторов, тогда как межгрупповые различия определяются еще и генетической изменчивостью.
Анализ изменчивости процесса обучения у гибридов от диаллельного скрещивания пяти инбредных линий мышей показал, что параметры выработки условной реакции избегания удара электрического тока в первый день обучения имеют наибольшую величину генетической изменчивости. На 5-й день обучения значительная вариабельность ответов наблюдается только внутри групп, из чего следует вывод о средовой природе этой изменчивости. Иными словами, наибольшие связанные с генотипом вариации поведения обнаруживаются на начальных стадиях обучения. Оценка размеров iipMF в этом эксперименте также показала существование отрицательной корреляции между величиной данного проекционного поля и быстротой усвоения навыка.
Эксперименты с тироксином. Если данная корреляция отражает причиннную зависимость между размером мозга и быстротой образования временной связи, то попытка изменить площадь iipMF путем воздействия на развивающийся мозг должна привести к изменениям в уровне обучения навыку избегания. Введение новорожденным крысятам тироксина ускоряло физическое развитие животных и развитие их поведения. В целом площадь iipMF проекций у взрослых крыс RHA, получавших тироксин, была большей и более изменчивой, чем у контрольных животных, а способность их к обучению обнаружила такую же корреляцию, как и в предыдущих экспериментах. Размер других структур гиппокампа у крыс после введения тироксина также варьировал, но корреляции с показателями обучения навыку избегания выявить не удалось. Аналогичные данные были получены при неонатальном введении тироксина и на мышах линий BALB/cn DBA (Lipp et al., 1989).
Исследовательская и локомоторная активность в "открытом поле". Следует отметить, что условная реакция активного избегания — это навык, с необходимостью формирования которого животные встречаются только в условиях эксперимента. В естественных и приближенных к естественным условиях существования животные, в том числе мыши и крысы, никогда не сталкиваются с ситуацией, когда, избежав опасности и попав в "безопасное" место, они снова слышат сигнал опасности и начинают воспринимать данную ситуацию как угрожающую. Иными словами, существование зависимости между обучением избеганию тока и площадью iipMF не дает достаточной информации для суждения о морфофизиологических механизмах адаптивных реакций. В связи с этим вызывал интерес характер зависимости других признаков поведения от величины площади iipMF.
Для ее изучения было выбрано исследовательское поведение в установке "открытое поле" мышей разных линий. Уровень исследовательской активности по ряду показателей оценивали в течение 20 мин теста в "открытом поле" (Crusio, 1995) с использованием схемы диаллельного скрещивания. У гибридов мышей инбредных линий можно было оценить не только фенотипические, но и генетические корреляции между признаками поведения и нейрогистологическими показателями. Полученные данные суммарно автор интерпретировал следующим образом. Мыши, имеющие большую по величине площадь проекции iipMF, всего за 20 мин достаточно эффективно собирают информацию о новой среде, в которую их поместили. И уровень страха (по интенсивности дефекации), и уровень исследовательской активности у них оказываются невысокими: обстановка теста быстро теряет для них новизну, и названные показатели поведения снижаются по ходу опыта сильнее, чем у мышей, с меньшей величиной iipMF. Действительно, у инбредных линий мышей, селектированных X.Ван-Абиленом на высокую и низкую частоту "стоек" в тесте "открытого поля" (см.: 8.6.2.1), В. Крузио нашел положительную корреляцию этого поведения с площадью iipMF.
В последующем были проанализированы другие признаки поведения, в частности обучение в так называемом радиальном лабиринте. Этот лабораторный тест был разработан американским исследователем Д.Олтоном (Olton, 1977) для исследования пространственной памяти животных.
Обучение животных в радиальном лабиринте считается тестом, который оценивает функции памяти, связанные с гиппокампом (Olton et ah, 1979). Успешность обучения в радиальном лабиринте мышей, различающихся по величине iipMF, положительно коррелировала с размером этой структуры (Schwegler, Lipp, 1995).
Другим тестом на способность к ориентации в пространстве и, следовательно, на уровень развития функции гиппокампа, считается ставший очень популярным тест в водном лабиринте Морриса (Morris, 1984). Этот тест предполагает, что мышь (крыса), плавая в непрозрачной воде небольшого бассейна, может научиться находить невидимую, скрытую под водой платформу, независимо от того, из какой точки периметра бассейна ее выпустили. Иными словами, животное обучается находить цель, ориентируясь в пространстве, не полагаясь на ее локальные признаки (Morris, 1984).
Успешность выполнения тестов на пространственную ориентацию зависит от функции гиппокампа. Оказалось, что выполнение теста Морриса, точнее, прочность сформированной пространственной памяти также положительно коррелирует с размером iipMF.
Иными словами, тесты на способность формировать представления о пространстве и о своем положении в нем животные усваивают тем успешнее, чем больше размер вариабельного поля проекции мшистых волокон гранулярных клеток на базальных дендритах пирамидных нейронов поля САЗ гиппокампа.
В большой серии работ, проведенных X.-П.Липпом, Г.Швеглером и их коллегами (Schwegler, Lipp, 1995), исследовали корреляции успешности выполнения "пространственных" и непространственных тестов с размерами не только iipMF, но и других областей гиппокампа. Однако такие фенотипические корреляции были обнаружены только для указанной области проекции мшистых волокон — нейронов зубчатой фасции. Был сделан вывод, что именно она играет важную роль в осуществлении и/или модуляции процессов обучения разных типов.
Существование описанных выше фенотипических корреляций не всегда может свидетельствовать о причинной связи между размером структуры и выраженностью функции. Однако в данном случае ее наличие подтверждается, во-первых, тем, что изменение размера iipMF с помощью тироксина (см.: выше) изменяет и поведение, а во-вторых, наличием не только феногипической, но и генетической корреляции между этими признаками (Crusio,1995).
Общим выводом из приведенного цикла работ по генетике поведения, существенно расширившего наши знания о роли гиппокампа в формировании навыков разной сложности, является следуюищи. Генетические системы, ответственные за формирование и ра* боту данного отдела мозга, и функциональные системы, "отвечающие" за обучение, совпадают или имеют общие звенья. В этих работах был детально разработан принципиально новый метод не-инвазивного контроля размера структур мозга. Этот размер можно было "варьировать" не путем частичного разрушения, что всегда влечет за собой много отрицательных последствий, а с помощью генетических методов. Корреляции размеров структуры с особенностями поведения, полученные таким путем, позволяют физиологам лать надежные выводы о физиологических механизмах последнего.
Исследование этой проблемы на генетических моделях (линиях мышей) продиктовано необходимостью выяснить механизмы нарушений строения мозолистого тела у человека.
Мозолистое тело (corpus callosum) представляет собой мощный проводящий путь (т.е. состоит из аксонов нейронов новой коры), функцией которого считается интеграция процессов возбуждения и торможения двух полушарий (подробнее см.: Шульговский, 1997; Lipp, Wahlsten, 1992).
Мозолистое тело соединяет между собой как гомотопические, т.е. зеркально расположеннные, так и гетеротопические зоны новой коры. В норме размер мозолистого тела, а следовательно, число волокон, связывающих два полушария, варьирует, причем выявить какие-либо корреляции с физиологическими функциями не удается.
Как в XIX в., так и в первой половине XX в. полное отсутствие мозолистого тела (агенез) у человека можно было обнаружить только post mortem (после смерти). С развитием метода компьютерной томографии оказалось возможным определять размеры мозговых структур и мозолистого тела, в частности, прижизненно. Это дало возможность оценить корреляции морфогенетического дефекта этой структуры с уровнем интеллектуального развития, выполнением вербальных тестов, функциональной асимметрией и другими психофизиологическими особенностями личности. Оказалось, что недоразвитие и/или полное отсутствие мозолистого тела не препятствует формированию нормальной речи (за исключением некоторых тонкостей, например способности к подысканию рифм). В то же время отмечено, что при недоразвитии мозолистого тела испытуемые имеют трудности с быстрым выполнением заданий, требующих двусторонней двигательной координации (Jeeves, 1986, цит, по: Lipp, Wahlsten, 1992). В целом у испытуемых с недоразвитием мозолистого тела коэффициент интеллектуальности IQ (см.: Равич-Щербо и др. 1988; Фогель, Мотульский, 1990) был несколько ниже среднего.
У крупных млекопитающих — лошадей, крупного рогатого скота, обезьян — недоразвитие мозолистого тела встречается редко, в отличие от лабораторной мыши, где это нарушение строения мозга наблюдается часто. У нескольких сублиний инбредной линии BALB/c, у линий 129/J, I/Ln и DDN недоразвитие или полное отсутствие мозолистого тела встречается в разном проценте случаев (Lipp, Whalsten, 1992). В частности, у мышей линии I/Ln и сублинии BALB/cWahl оно полностью отсутствует почти у 100% животных. Не обнаружено никаких физиологических или поведенческих особенностей, по которым можно было бы определить степень развития этой структуры у мыши, кроме прямого анатомического исследования мозга.
Установлено, что причиной недоразвития или отсутствия мозолистого тела в мозге мыши является особенность процесса формирования межполушарных аксонных связей в период внутриутробного развития. Причиной возникновения дефекта является, по-видимому, задержка процесса роста аксонов, образующих мозолистое тело. В результате аксоны не могут преодолеть препятствие в виде тяжа глиальных клеток, расположенного у них на пути. Часть таких волокон все-таки прорастает в противоположное полушарие, иногда "используя" для этого гиппокампову коммиссуру. Другая часть растет в том же полушарии по его продольной оси, образуя так называемый пучок Пробста (по имени ученого, его впервые описавшего — Probst, 1901, цит. по: Lipp, Wahlsten, 1992).
В какой степени мозг новорожденной мыши будет аномальным, зависит от ряда факторов, одним из которых являются условия питания плода в организме матери. Оказалось, что если в процессе развития эмбрионов самка мыши еще продолжает выкармливать молоком предыдущий помет, то вероятность недоразвития мозолистого тела у детенышей нового помета оказывается выше (Wahlsten, 1982).
Генетические исследования линий, подверженных этому анатомическому дефекту, показали, что недоразвитие мозолистого тела определяется, по-видимому, только двумя генами с аутосомной локализацией.
Для оценки влияния недоразвития мозолистого тела на поведение было проведено сравнение линий животных с нормальным мозолистым телом и с его полным отсутствием (агенезом). Очевидно, что обнаруженные межлинейные различия можно будет связать с различиями в строении мозолистого тела лишь предположительно, поскольку инбредные линии различаются между собой по очень большому числу локусов, каждый из которых потенциально также может определять особенности поведения. В связи с этим предполагаемую роль мозолистого тела необходимо проанализировать дополнительно. Было исследовано поведение мышей линий BALB/c, 129/J или DDN, особи которых варьируют по степени развития мозолистого тела, а также поведение потомков от скрещивания двух линий с изменчивостью этого морфологического признака (Lipp, Wahlsten, 1992).
Для анализа поведения таких гибридов было выбрано предпочтение одной из лап в пищевой пробе. В таком тесте в узкую трубку помещают маленький кусочек пищи, который мышь может достать только одной лапой. Устойчивость в предпочтении правой или левой лапы часто оказывается четкой индивидуальной характеристикой животного. Оказалось, что недоразвитие или полное отсутствие мозолистого тела отрицательно влияет на склонность животного пользоваться только одной лапой (Lipp, Walilsten, 1992). Мыши с недоразвитием и полным отсутствием мозолистого тела достоверно слабее выполняли специальный тест на координацию движений задних конечностей. Обучение навыку побежки в К-образном лабиринте, где животные обучаются побежкам направо или налево в зависимости от текстуры внутренних стенок лабиринта, у мышей 129/J и, в особенности, у I/Ln сильно нарушено. В то же время у них не отмечается особых дефектов при обучении реакции активного избегания в челночной камере. В тесте водного лабиринта Морриса формирование пространственного навыка отыскания ''невидимой'' платформы также не показывает заметной корреляции с размером мозолистого тела, исключение составляли мыши линии I/Ln, которые медленно выполняли тест из-за сильной тенденции к плаванию малыми кругами.
Поведение мышей с недоразвитием мозолистого тела изучено еще недостаточно, в частности неясно, в какой степени у них изменена латерализация*) функций, отражающая специфику физиологии больших полушарий.
*)* Латерализация, т.е. специализация функций правого и левого полушарий, привлекает все большее внимание исследователей и обнаруживается не только у человека (Спрингер, Дейч,. 1977), но и у многих животных (Бианки, Филиппова, 1987).
Линии мышей с разной степенью дефекта развития мозолистого тела представляют собой уникальную биологическую модель для оценки поведения у генетически идентичных животных и выявления особенностей поведения, порождаемых именно данным нейроанатомическим дефектом.
Как мы уже видели, существует генетически детерминированная изменчивость размеров некоторых структур мозга. Оказалось, что успешность выработки двусторонней реакции активного избегания в челночной камере положительно коррелировала не только с общим объемом гиппокампа, но и с числом гранулярных клеток зубчатой фасции (Wimer et al., 1983).
Используя подсчеты числа нейронов в ряде полей гиппокампа и методы классического генетического анализа, Ц. и Р.Ваймеры (С.Wimer, R.Wimer, 1982, 1989) действительно, показали, что количество нейронов основных полей гиппокампа определяется небольшим числом генов (от 2 до 5). В дальнейшем межлинейные различия в числе нейронов были обнаружены и в других исследованиях. Число дофаминергических нейронов в вентральном отделе ствола у линии СBА оказал ось меньше, чем у улинии BALB/c. Такие же межлинейные различия были найдены в гипоталамусе. Сравнение других линий показало, что в голубом пятне у линии BALB/c катехоламинергических клеток на 38% меньше, чем у линии C57BL. По данным другой работы, у мышей линии DBA/2 в медиовентральном гипоталамусе имеется клеточное скопление — так называемая pars compacta, тогда как у мышей линии C57BL/6 оно отсутствует, а у самок DBA/2 его объем на 80% больше, чем у самцов. Количество нейронов в этих отделах коррелирует с некоторыми показателями исследовательской активности (см.: Полетаева, Романова, 1990).
Возможно, что одна из причин изменчивости числа клеток взрослого мозга — различия в числе клонов, формирующих данный участок мозга, и/или в числе митотических делений исходной клетки перед началом дифференцировки. Еше одной причиной генетически обусловленных различий в числе клеточных элементов нервной системы могут быть вариации в действии генетически контролируемой системы апоптоза — так называемой "программированной гибели клеток" в процессе нейрогенеза (см.: Finlay et al., 1987).
В разделе 8.2.5 были частично рассмотрены возможные механизмы реализации изменчивости поведения. Одним из них могут быть межвидовые (при сравнении близких видов), межлинейные и/или межпопуляционные различия в порогах проявления дискретных поведенческих актов ФКД. Межлинейные различия в числе стоек или норковых реакций или же, например, в частоте проявления таких видоспецифических реакций, как подтаскивание детенышей к гнезду (Hurnick, 1973), или в движениях при постройке гнезда (Lynch, 1980) и др., могут быть отражением существования разных порогов проявления этих движений. Эти различия, по крайней мере частично, могут быть связаны и с разным числом нервных элементов в важной для выполнения данного поведения мозговой структуре.
Несомненная связь между числом нервных элементов в мозге и сложностью поведения существует на уровне межвидовых сравнений и сравнений между более крупными таксономическими единицами, однако можно предположить существование и более тонкой зависимости на внутривидовом уровне.
Высокие темпы эволюции мозга млекопитающих и формирование мозга человека традиционно до сих пор не имеют единого объяснения с точки зрения современной теории эволюции. С.Н. Давиденков (см.: 2.5.5), анализируя генетические основы функций мозга, предположил, что чем меньше филогенетический возраст данной функции, тем более вариабельна она оказывается. Сходные с этим, но более развернутые предположения делают и современные исследователи (см.: Lipp, 1989; Lipp, Wolfer, 1995).
Известно, что с позиций эволюционной теории большая продолжительность жизни и малое число потомков животных (млекопитающих) крупных видов, представляют собой в известном смысле помехи процессу эволюции. Эволюция таких таксономических групп животных должна была бы идти более медленно, однако этого не происходит, и скорость эволюции видов, имеющих крупные размеры и сложно устроенный мозг, примерно такая же, как и видов более мелких, с более простым мозгом. Отсюда, совершенствование высших психических функций, можно рассматривать как биологический механизм компенсации таких помех.
Проф. Х.-П. Липп высказывает гипотезу о существовании особой категории эволюционных морфогенетических процессов, которые способствуют формированию сложного субстрата психических способностей.
Высшие (по сложности функции) системы мозга в процессе индивидуального развития дифференцируются в наиболее поздние сроки. Х.-П.Липп предполагает, что некоторые мутации, вызывающие изменения структуры высших отделов мозга, защищены от элиминирующего действия естественного отбора благодаря существованию так называемых буферных систем. Под буферными системами автор понимает совокупность генетически детерминированных физиолого-биохимических механизмов, обеспечивающих гомеостаз развития, способность мозговых структур к реорганизации в процессе развития, процессы, обеспечивающие пластичность взрослого мозга. Действие этих буферных систем обеспечивает стабильность функционирования организма и "перекрывает" возможные функциональные последствия слабых мутаций, которые в отсутствие этих "буферов" могли бы влиять на поведение.
Следующее предположение заключается в том, что накопление генетической изменчивости высших структур мозга все же обнаруживается в виде появления некоторых структурных изменений. Это, в свою очередь, может быть причиной возможной реорганизации мозга в направлении "сверху вниз", т.е. может повлечь за собой реорганизацию и более низко расположеннных отделов ЦНС.
Предполагается, что в геноме видов, имеющих сложный мозг, существуют группы исключительно изменчивых генов (названных "вариобоксом"), которые и "ответственны" за индивидуальные различия способностей, темперамента и др.
Эта концепция имеет эвристическую ценность для выявления связи между генами, мозгом и поведением. Она представляет собой теоретическую канву, которая необходима для планомерной оценки новой информации о поведении мышей с искусственными мутациями и мышей-нокаутов.
При написании этого раздела мы использовали многочисленные примеры как из оригинальных публикаций, так и из обобщающих монографий, учебных пособий и сборников. Для более подробного знакомства с предметом можно рекомендовать следующие книги.
Руководство по физиологии (том "Генетика поведения и физиологических признаков", 1981), учебники Л.Эрман и П.Парсонса (1984), Д.Хэя (Hay, 1985), а также прекрасные методические пособия D.Goldowitz et al. (eds.) "Techniques for the genetic analysis of brain and behavior. Focus on the mouse" (1992) и P.Driscoll (ed.) "Genetically defined animal models of neurobehavioral disfunction" (1992).
Развитие науки идет стремительными темпами. К этому трудно привыкнуть, и еще труднее, даже вооружившись современными методами "слежения" за ее развитием (такими как Internet, Medline, Science Citation Index и пр.), успевать реагировать на эти изменения при издании учебника.
Однако в биологии существует и мощная "консервативная" составляющая. Наряду с эволюционным учением, зоологией, ботаникой, экологией, генетикой и цитологией, биохимией и физиологией, ее основу составляет и этология — наука о поведении животных.
Именно поэтому авторы надеются, что несмотря на бурный прогресс молекулярных основ нейрогенетики и нейрофизиологии написанное в этой книге окажется необходимым и полезным начинающим биологам.
Авторы также надеются, что эта книга поможет ее читателям осознать, что наши современные знания — это в первую очередь заслуга наших предшественников и учителей, которым мы все должны быть бесконечно благодарны.