
НЕЙРОБИОЛОГИЯ
Природа, 1997, № 3, стр. 81-89.
Нейробиология индивидуальности
П. В. Симонов
Павел Васильевич Симонов, нейрофизиолог, академик РАН, директор Института высшей нервной деятельности и нейрофизиологии, академик-секретарь Отделения физиологии. Область научных интересов — нейрофизиология и психофизиология мотиваций и эмоций.
ПОИСКИ естественнонаучных основ индивидуальных особенностей психики и поведения продолжаются не одно столетие. Античные авторы полагали, что психический склад человека определяется соотношением четырех жидкостных сред организма: крови, желчи, черной желчи и лимфы. Греческие названия этих жидкостей дали имя соответствующим темпераментам: сангвиническому, холерическому, меланхолическому и флегматическому.
Поистине революционным моментом в истории изучения естественнонаучных основ индивидуальных различий стала концепция И.П.Павлова о типах нервной системы человека и животных. Поскольку высшая нервная (психическая) деятельность, определяющая внешне реализуемое поведение, есть функция головного мозга, именно в деталях деятельности мозга следует, по мысли Павлова, искать причину индивидуальных различий психики и поведений. Основывая свою классификацию на свойствах возбуждения и торможения нервных клеток, Павлов понимал, что путь от элементарных нервных процессов к внешне реализуемому поведению лежит через взаимодействие макроструктур — функционально специализированных отделов головного мозга.
Хотя вопрос «Что такое эмоция?» — Уильям Джеймс сделал заглавием своей статьи, опубликованной более 100 лет назад [1], он и сегодня остается без общепризнанного ответа. Вот что мы можем прочитать в руководстве «Физиология человека», вышедшем из печати через сто лет после статьи Джеймса: «Несмотря на то, что каждый из нас знает, что такое эмоция, дать этому состоянию точное научное определение невозможно... В настоящее время не существует единой общепризнанной научной теории эмоций, а также точных данных о том, в каких центрах и каким образом эти эмоции возникают и каков их нервный субстрат» [2].
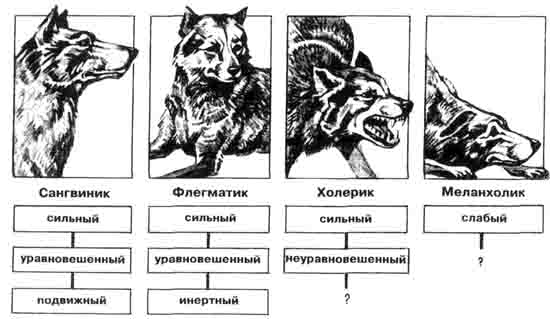
Типологическая классификация Гиппократа.
Тридцать лет тому назад результаты психофизиологических экспериментов привели меня к заключению о том, что эмоции высших животных и человека определяются какой-либо актуальной в данный момент, витальной (голодом, жаждой и т.п.), социальной или идеальной (духовной) потребностью и оценкой вероятности ее удовлетворения на основе фило- и ранее приобретенного онтогенетического опыта. Эту оценку субъект выполняет непроизвольно и зачастую неосознанно, сопоставляя информацию о средствах и времени, предположительно необходимых для удовлетворения потребности, с информацией, которой он располагает в данный момент. Низкая вероятность достижения цели ведет к отрицательным эмоциям (страху, тревоге, гневу, горю и т.п.), которые субъект активно стремится свести к минимуму. Возросшая вероятность удовлетворить потребности по сравнению с ранее имевшимся прогнозом порождает положительные эмоции удовольствия, радости, торжества, которые субъект стремится усилить, продлить, повторить. Термин «информация» я употребляю, имея в виду его прагматическое значение, определяемое изменением вероятности достижения цели [3]. назвали «общим законом человеческих эмоций».
В 1984 г. Д.Прайс и Дж.Баррел воспроизвели наши опыты в чисто психологическом варианте, предложив испытуемым мысленно представить себе какое-либо эмоционально окрашенное событие и затем отметить на специальных шкалах силу своего желания, предполагаемую вероятность достижения цели и степень переживания [4]. Количественная обработка полученных данных подтвердила существование зависимости, которую авторы
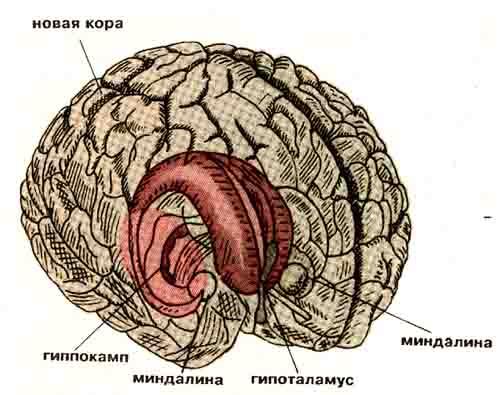
Структуры мозга, участвующие в формировании эмоциональных реакций.
Многочисленные эксперименты с повреждением отдельных образований мозга и последующей регистрацией их электрической активности привели к выводу о том, что среди структур, участвующих в генезе эмоциональных реакций, можно выделить две системы. Одна из них (будем называть ее информационной) оценивает вероятность удовлетворения потребности, а другая (мотивационная) выделяет доминирующую потребность, требующую первоочередного удовлетворения. К первой системе относятся фронтальные (лобные) отделы новой коры мозга и гиппокамп — участок древней коры, напоминающий по форме морского конька. Ко второй — ядра амигдалы, названные так за сходство их очертаний с миндалем, и расположенный в глубине мозга под зрительным бугром гипоталамус. В результате сложного взаимодействия этих четырех мозговых образований формируются вектор поведения и соответствующее ему эмоциональное состояние, переживаемое человеком или животным в данный момент. Позднее к аналогичным выводам пришли и другие авторы [5].
Дальнейшие исследования позволили предположить, что индивидуальные особенности функционирования, а точнее относительная активность каких-либо из перечисленных четырех отделов мозга, лежат в основе античных темпераментов, типов нервной системы Павлова и параметров Г.Айзенка [6]. По мнению К.Клонингера, темперамент больше связан с миндалиной и гипоталамусом, в то время как характер определяется функционированием гиппокампа и новой коры [7].
В работе, о которой пойдет речь, мы провели нейрофизиологический анализ двух форм поведения, достаточно стабильно характеризующих подопытное животное. Это — способность к так называемому «самоконтролю», к задержке произвольной двигательной реакции ради получения более ценной награды и способность к выработке условной реакции избегания, предотвращающей болевое раздражение другой особи того же вида — феномен «эмоционального резонанса».
МОЗГОВЫЕ МЕХАНИЗМЫ СВОБОДНОГО ВЫБОРА
Психологи сравнительно давно описали эксперименты, в которых детям предлагалось большее или меньшее вознаграждение сладостями в зависимости от того, хватали ли они их сразу же после ухода экспериментатора из комнаты или, согласно инструкции, терпеливо ожидали его возвращения. Оказалось, что этот тест имеет важное прогностическое значение. Те, кто в четырехлетнем возрасте выдерживал испытание, росли более уживчивыми, общительными, изобретательными и уверенными в себе. Поддавшиеся соблазну вырастали нерешительными, одинокими, упрямыми, уязвимыми к стрессовым воздействиям.
Мы воспроизвели похожие эксперименты в опытах на кошках [8]. Условный сигнал — свет, который загорался под прозрачной крышкой педали. Если кошка нажимала на педаль в течение 2 с, она получала менее привлекательную •для нее смесь хлеба с 30%-м содержанием мяса. Если же она игнорировала первый сигнал и, выждав 7 с, нажимала на повторно засветившуюся педаль, ее ожидало лакомое мясо. Под наркозом кошкам вживляли во фронтальную кору и латеральное ядро гипоталамуса пучки из четырех—пяти нихромовых электродов диаметром 50 мкм каждый. Данные электрической активности нервных клеток обрабатывали по специальной программе для разделения нейронных спайков (это — импульсная часть потенциала действия нейрона) по форме, по накоплению импульсных рядов из спайков одинаковой формы. Временные (по 15 с) фрагменты записей мультинейрон-ной активности анализировали одновременно из фронтальной коры и латерального ядра гипоталамуса непосредственно перед сигналом и сортировали их в зависимости от выполнения коротко-или длинноотставленной пищедобыва-тельной реакции.
Из четырех кошек (самцов), взятых в опыты, две предпочитали получать мясо, т.е. использовали стратегию выжидания, а две не выдерживали отсрочки и довольствовались мясосу-харной смесью. В специальных опытах после 24-часового голодания кошки не изменили поведения, характерного для каждого из них.
При оценке взаимодействия исследуемых структур мозга мы руководствовались следующим принципом: когда нейронные спайки в гипоталамусе опережали таковые во фронтальной коре, мы говорили о гипоталамо-фрон-тальном взаимодействии; а при обратной последовательности спайков — о фронтально-гипоталамическом. Оказалось, что у «импульсивных» кошек и «контролирующих» поведение фронтально-гипоталамические взаимодействия были одинаковые. Иное дело — гипоталамо-фронтальные связи: у кошек, способных к «самоконтролю», преобладают взаимодействия с длинными временными задержками, а у импульсивных наблюдаются короткоот-ставленные. Это правило справедливо и для тех случаев, когда кошки в отдельных опытах проявляли не типичную для них реакцию. Что касается взаимодействия между близко расположенными нейронами внутри фронтальной коры, то они не обнаружили связи с типологическими особенностями исследованных животных.
В другой серии экспериментов шести кошкам (самцам) электроды вживляли во фронтальную и моторную кору — в зону представительства передней лапы, нажимающей на педаль9. В этом случае фронтально-моторные межнейронные взаимодействия с временными задержками до 100 мс не отличались у кошек, работающих импульсивно или в режиме «самокон--троля», т.е. не определяли типологических особенностей животных. Межнейронные связи с задержками до 30 мс начинали преобладать у кошек обоих групп при реализации короткоотстав-ленных реакций. Число фронтально-моторных нейронных связей в диапазоне О—30 мс увеличивалось у трех необученных кошек после суточного голодания [10].
Наконец, из четырех кошек, которым электроды вживлялись в префрон-тальную кору (поле 8) и базолатераль-ную миндалину, у трех наблюдали тенденцию к «самоконтролю», а четвертая оказалась «импульсивной». В ситуации активного выбора задержки двигательной реакции, продолжительность которой определяла ценность подкрепления, количество внутриамиг-далярных и фронтально-амигдалярных нейронных взаимодействий преобладало у «импульсивной» кошки по сравнению с животными, способными к «самоконтролю». Главное же состоит в том, что наибольшая активация нейронных взаимодействий с участием миндалины зарегистрирована у всех кошек при выпадении двигательной реакции, т.е. при затруднении выбора животным стратегии своего поведения.
Незавершенность экспериментов по выяснению роли миндалины и отсутствие данных относительно участия гиппокампа не позволяют нам представить достаточно полную картину событий, разыгрывающихся в нейронах головного мозга в ситуации свободного и потому трудного выбора между «синицей в руках» и «журавлем в небе». Пока можно высказать только несколько предварительных замечаний.
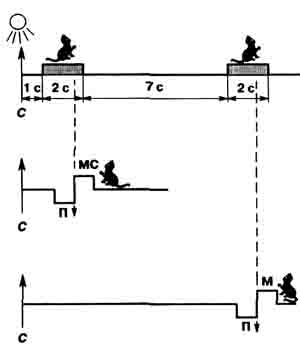
Схема эксперимента по выработке условной реакции избегания у кошек. Вверху — период эффективной педали, время в с; в середине ~ ' короткоотставленная реакция, мс — мясосу-харная смесь; внизу — длинноотставленная реакция, м — мясо.
Судя по полученным результатам, решающую роль в реализации выбора поведения с более или менее ценным подкреплением играет влияние мотивационных структур гипоталамуса на передние отделы новой коры, но отнюдь не обратное влияние «интеллектуальных» структур коры на мотива-ционную сферу. Как тут ни вспомнить гениальное утверждение Бенедикта Спинозы: страсти побеждаются не разумом, но более сильными страстями. Что касается миндалины, то она вовлекается в деятельность тем сильнее, чем труднее выбор между конкурирующими «страстями». Судя по другим нашим экспериментам, в этой конкуренции участвуют эмоции, оценивающие не только силу и качество мотиваций, но и вероятность их удовлетворения при той или иной стратегии поведения [11].
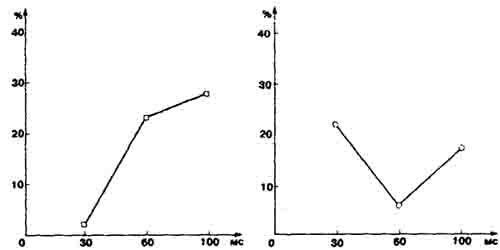
Количество (%) межнейронных гипоталамо-фронтальных связей у кошек, способных к «самоконтролю» (цветная кривая) и «импульсивных».
НЕЙРОНЫ СОЧУВСТВИЯ
В предисловии к последнему прижизненному изданию своего классического труда «Интегративная деятельность нервной системы» Чарльз Шеррингтон писал: «Лорд Актон намеревался создать «Историю свободы», между тем не менее стоящим было бы создание «Истории альтруизма». Это ,м,ожет быть сочтено отходом от физиологии, однако я не думаю, что это так... В той мере, в какой физиология включает в себя человека как физиологический фактор на нашей планете, это противоречие, главным действующим лицом которого он является, не лежит вне границ физиологической науки»12.
В своих исследованиях, идущих на протяжении последних 20 лет, мы решили начать «историю альтруизма» с... белых беспородных крыс, использовав характерное для этих животных предпочтение находиться в ограниченном пространстве [13]. В специальной установке пол плексигласового «домика» представлял педаль, которая автомагически включала счетчик времени и болевое раздражение электрическим током лап другой крысы, находившейся за тонкой прозрачной звукопроницаемой перегородкой. Подопытное животное один раз в день помещали в открытую, относительно просторную часть установки на 5 мин и регистрировали время его пребывания на педали. В течение 5 дней вход в «домик» не сопровождался болевым раздражением второй крысы, в то время как на протяжении следующих 10 дней при каждом появлении подопытного животного в «домике» включали ток силой 1—2 мА. Раздражение партнера продолжалось 3—5 с с пятисекундными интервалами до тех пор пока исследуемая крыса находилась на педали.
Из 247 крыс (самцов) 77 особей (т.е. 31%) сравнительно быстро выработали условную реакцию избегания болевого раздражения партнера; 111 (45%) пришли к тому же, только побывав несколько раз в качестве «жертвы»; а у 59 крыс (24%) вообще не удалось выработать условный рефлекс избегания сигналов оборонительного возбуждения партнера (крика, подпрыгивания, выделения специфических пахучих веществ). Крыс первой группы мы в дальнейшем будем называть альтруистами, а последней — эгоистами.
Нобелевский лауреат Г.Саймон считает, что альтруизм существует до тех пор, пока выгоды от него для вида превышают потери от уменьшения индивидуальной приспособленности [14]. Может быть, именно поэтому распределение особей, чувствительных и нечувствительных к сигналам боли, исходящим от партнера, оказалось примерно одинаковым среди крыс, собак, обезьян и... людей [15], будучи подчинено соотношению 1:2.
Локальное повреждение структур головного мозга у крыс позволило выявить: мозговые образования, повреждение которых не оказывает достоверного влияния на реакцию избегания (например, гиппокамп); структуру, эффект повреждения которой отчетлив и вместе с тем не зависит от индивидуальных особенностей животного (фронтальные отделы новой коры); отделы мозга, последствия повреждения которых определяются индивидуальными особенностями животного до операции (ядра амигдалы и гипоталамус). Именно поэтому в качестве объекта для нейрофизиологи-ческого анализа мы выбрали гипоталамус.
В первой серии экспериментов стояла задача исследовать импульсную активность нейронов эмоционально позитивных структур латерального гипоталамуса и латеральной преоптической области (электрическая стимуляция этих образований вызывала реакцию самораздражения) и эмоционально негативной области вентромедиального гипоталамуса при внутримозговых эмоциональных воздействиях разного знака [16]. Негативное «наказующее» воздействие достигалось стимуляцией дорзомедиального тегментума, которая вызывала реакцию избегания того места, где применялось болевое раздражение. Нейронную активность регистрировали у 58 бодрствующих, свободноподвижных беспородных крыс (самцов). Затем анализировали 30-секундные отрезки записи нейронной активности, усредненные по 5 или 10 наложениям, до и после электрической стимуляции эмоционально позитивных и негативных структур. Для статистической обработки данных использовался непараметрический критерий Вил коксона.
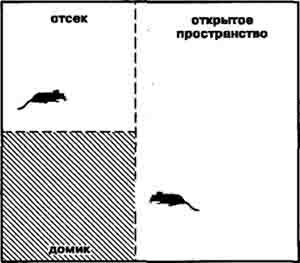
Экспериментальная камера отбора альтруистов и эгоистов среди крыс,
Выяснилось, что в эмоциогенных структурах латерального гипоталамуса, латерально преоптической вентромеди-альной области имеются нейроны, которые реагируют на эмоционально позитивные и негативные внутримозго-вые стимулы разнонаправленно (возбуждением или торможением). В ответах нейронов вентромедиального гипоталамуса на внутримозговые и периферические (болевое раздражение хвоста) эмоционально негативные воздействия преобладало торможение. В структурах латерального гипоталамуса и латеральной преоптической области количество тормозных и активационных ответов было приблизительно одинаково. Кроме того, в исследованных структурах гипоталамуса обнаружили 4% нейронов, отвечавших резкой активацией на отсутствие ожидавшихся крысой эмоционально негативных воздействий. О существовании в латеральном гипоталамусе нейронов, противоположным образом реагирующих на эмоционально позитивные и негативные стимулы, сообщается в работе Т. Оно и К.Накамура [17].
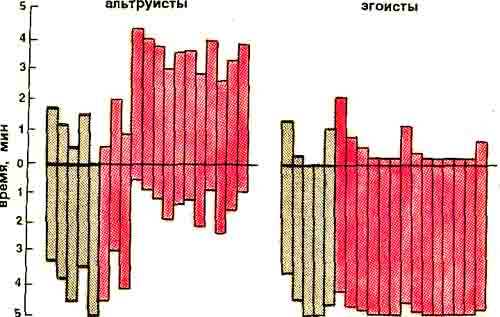
Выработка реенкцю избегания у крысы-альтруиста и у крысы-эгоиста. Ниже горизонталь -&QU линии - время требьюа-'шш на шедалм, выше - в открытом пространстве в минутах.. Серые- столбцы - до раздраженная током лв.рт-аере, цветные - при раздражении. По горизонтали - дни экспериментов.
Теперь мы могли перейти к решению основной задачи: ответить на вопрос о том, как реагируют нейроны гипоталамуса на сигналы оборонительного возбуждения другой особи того же вида18. Отметим, что такого рода данных мы в существующей литературе не нашли.
С помощью описанной методики выработки реакции избегания болевого раздражения партнера среди 20 крыс (самцов) выявили 8 альтруистов, чье время пребывания в «домике» не превышало 100 с, и 8 эгоистов, которые, не обращая внимания на крики партнера, большую часть времени проводили на педали. Затем для регистрации нейронной активности им вживляли семиствольные нихромовые электроды диаметром 50 м км в латеральный гипоталамус и латеральную преоптическую область и электроды диаметром 100 мкм для электрической стимуляции этих структур. В дорзомедиальный тегментум вживляли только стимулирующие электроды Регистрировали либо одиночные нейроны, либо из мультиклеточ-ной активности выделяли два-три ряда спайков, различающихся по амплитуде не менее чем на 30%. Данные обрабатывали с помощью критерия Стьюдента (различия считали значимыми при р меньше 0,05). Исследовали влияние на импульсную активность нейронов крика крысы-партнера, раздражаемой электрическим током (1—2 мА), и внутримозговой стимуляции эмоционально позитивных (латерального гипоталамуса и латеральной преоптической области) и эмоционально негативных (дорзомеди-ального тегментума) структур.
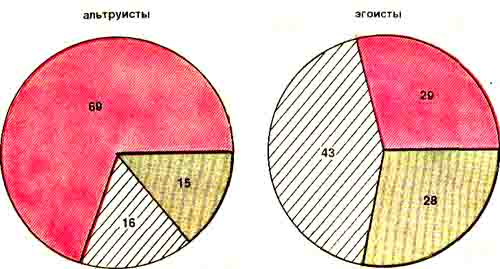
Оказалось, что у крыс-альтруистов доминируют (69%) нейроны, ответы которых на крик сходны с реакцией на эмоционально негативную стимуляцию. У крыс-эгоистов, напротив, обнаружилась тенденция к увеличению (до 43%) ответов, аналогичных реакциям на эмоционально позитивные стимулы.
Полученные факты дают основание считать, что восприятие сигналов оборонительного возбуждения партнера создает у наблюдателя-альтруиста эмоционально негативное состояние (феномен «эмоционального резонанса»), которое и побуждает крысу прервать или даже предотвратить болевое раздражение «жертвы». Судя по полученным данным, этот механизм ослаблен у крыс-эгоистов, более того, до 43% зарегистрированных у них нейронов реагируют на крик «жертвы» как на эмоционально позитивную стимуляцию. Вряд ли наших эгоистов можно заподозрить в склонности к садизму. Скорее этот парадоксальный факт объясняется тем, что сигналы неблагополучия партнера крыса-наблюдатель воспринимает в ситуации, где, как она быстро убеждается, ей самой ничего не грозит.
Результаты выполненного иссления подтверждают, что в основе типологических различий поведения высших животных и, по-видимому, человека лежат индивидуальные особенности взаимодействия функционально специализированных макроструктур головного мозга. Решающее слово здесь будет принадлежать использованию новейших технологий прижизненного картирования функций мозга: позитронно-эмиссионной томографии, функционального магнитного резонанса, электрофизиологического картирования.
С другой стороны, изучение естественнонаучных основ индивидуальных различий представляет несомненный интерес для научно обоснованной педагогики. Эксперименты с повышением чувствительности крыс к сигналам неблагополучия партнера после собственного ознакомления с аверсив-ным (негативным) воздействием побуждают вспомнить призыв выдающегося педагога В.А.Сухомлинского о необходимости целеустремленно развивать у детей склонность к состраданию, сопереживанию и сочувствию. Воспитание не «мечтательной, а деятельной любви» (Ф.М.Достоевский), радость от успеха действий, направленных на облегчение участи другого, — путь к формированию нравственно полноценной личности. Самый лучший человек тот, говорил Л.Н.Толстой, кто живет преимущественно своими мыслями и чужими чувствами, а самый худший сорт человека — который живет чужими мыслями и своими чувствами... Из возможных сочетаний этих четырех основ, мотивов деятельности — все различие людей.
1. James W. What is an emotion? // Mind. 1884. V.9. № 34. P.188.
2. Физиология человека. Т.Ч. М., 1985. С.211.
3 Симонов П.В. Что такое эмоция? М., 1966; Симонов П.В. Эмоциональный мозг. М., 1981.
4 Price D., Barrell J. Some general laws of human emotion: Interrelationship between intensites of desire, expectation and emotional feeling // J.Person. 1984. V.52. P.389.
5. Escobar A. Nuevos conceptos sobre la significacion morfo functional del sistema limbico // BoI.estud.med. у biol. 1986. V.34. № 1. P.25-34; Do ux Le J. Cognitive-emotional interaction in the brain // Cognition and Emotion. 1989. V.3. № 4. P. 267-289.
6. Si mono v P.V. Individual characteristics of brain limbic structures as the basis of Pavlovian-Eysenckin thypology // Personality dimensions and arousal. N.Y„ 1987. P.121-132; Simonov P.V. The Motivated Brain. Philadelphia, 1991. P.280. ^Cloninger C. Temperament and personality // Current Opinion in Neurobiology. 1994. V.4. № 2. P. 266-273.
8. Мержанова Г.Х., Берг А.И., Мартинсон Ю.Л. Фронтально-гипоталамическое взаимодействие у кошек при реализации свободного выбора подкрепления // Журн. высш. нерв. деятельности. 1993. Т.43. № 3. С.487-495.
9. Мержанова Г.Х., Берг А. И. Фронтально-моторные межнейронные взаимодействия у кошек при реализации свободного выбора подкрепления // Там же. 1994. Т.44. № 6. С.954-962.
10. Мержанова Г.Х., Долбакян Э.Е. Временная организация межнейронных фронтально-моторных отношений в неокортексе кошек в зависимости от уровня пищевой мотивации // Там же. 1996. Т.46. № 3. С.97-104.
11. Si mono v P.V. The Emotional Brain. N.Y.; London, 1986. P. 267.
12. Шеррингтон Ч. Интегративная деятельность нервной системы. Л., 1969. С. 26.
13. Si mono v P.V. Conditioned avoidance responses to the pain stimulation of another animal // Activitas nervosa superior. 1977. V.19. № 1. P. 28-29.
14. Саймон Г. Что такое альтруизм? // В мире науки. 1991. № 5. С. 59.; Индивидуальный мозг / Под редакцией П.В.Симонова. М., 1993. С. 127.
15. Milgram S. Obedience to authority. N.Y., 1974.
16. Зайченко М.И., Михайлова Н.Г., Райго-р о деки и Ю.В. Реакции нейронов эмоциональных зон гипоталамуса при эмоциональных воздействиях разного знака // Журн. высш. нервн. деятельности. 1995. Т.45. Ns 2. С.367-376.
17. О по Т., Nakamura К. Learning and integration of rewarding and aversive stimuli in the rat lateral hypothalamus // Brain Research. 1985. V.346. P.368.
18. Зайченко М.И., Михайлова Н.Г, Райгородский Ю.В. Нейроны гипоталамуса и зоосоциальные взаимодействия // Журн. высш. нервн. деятельности. 1996. Т. 46. № 3. С.128-135.
© П.В.Симонов
(Последние исправления - 20.04.2000)