В этой последней части, состоящей из трех глав. мы рассмотрим весьма спорный вопрос о том, существует ли какое-либо сходство между психической деятельностью человека и животных. В гл. 26 рассматривается проблема языка и психических представлений, в том числе вопросы о том, способны ли обезьяны овладеть языком и могут ли животные иметь какие-либо намерения. Гл. 27 посвящена проблемам интеллекта животных, использования ими орудий (иногда рассматриваемого как признак интеллекта) и культурной эволюции. В гл. 28 мы обсудим вопросы, связанные с элементами самосознания и эмоциями у животных. Мы исследуем вопросы сознания и страдания животных с точки зрения их благосостояния. И наконец, мы обсудим проблему страдания животных с точки зрения эволюции.
Эдвард Толмен (1886-1959)
Эдвард Толмен, американский психолог, был «колючкой в теле современного ему бихевиоризма». Во многих отношениях его мышление опережало время. Можно сказать, что он был отцом современного когнитивного подхода к пониманию поведения животных. В отличие от представлений других когнитивных теоретиков своего времени, таких, как Джордж Романее (Romanes) или Вольфганг Кёлер (Kohler), представления Толмена не были менталистическими. Он считал поведение целенаправленным, но система его представлений не была антропоморфичной. Он был убежден, что животные ведут себя целенаправленно, но он никогда не считал, что у них имеется психический образ цели. Толмен признавал себя бихевиористом, но разрабатывал идею молярного, а не молекулярного бихевиоризма. С точки зрения молярного бихевиоризма, поведенческий акт как таковой имеет свои, присущие только ему характерные особенности, которые можно описать независимо от конкретных физиологических процессов, обеспечивающих данное поведение. Что касается молекулярного бихевиоризма, то он является редукционистским в том смысле, что стремится объяснить поведение через лежащие в его основе физиологические и физические явления. Основной работой Толмена была книга «Целенаправленное поведение животных и человека» (1932), однако позже он опубликовал много статей, в которых критиковал современные ему взгляды, опираясь на теоретические аргументы и экспериментальные данные, а также уточнял свои когнитивные представления.
Когнитивный подход Толмена (1932) к пониманию условного рефлекса Павлова имеет много общего с современным пониманием этого вопроса (Rescoria, 1978). Например, он считал, что животное научается, познавая подкрепляющий фактор, а не просто научается потому, что получает подкрепление. Он критиковал общепринятую теоретическую концепцию стимул - реакция и первым высказал мысль о том, что условный раздражитель - это сигнал, свидетельствующий о следовании за ним какого-то другого события. Толмен также был первым, кто выдвинул идею когнитивных карт. По его мнению, животное приобретает когниции, или знания обо всех деталях ситуации, которые организуются таким образом, чтобы их можно было использовать, когда они понадобятся. Полемизируя с господствовавшим в то время мнением о том, что животные «научаются посредством действий», Толмен показал в своих экспериментах, что животные могут получить информацию об общих характеристиках экспериментальной камеры или лабиринта, не осуществляя соответствующего поведения. Исследования Толмена привели к предположению, что во время научения животное приобретает когнитивную карту, указывающую, каким образом соответствующие причинные или пространственные характеристики внешнего мира связаны друг с другом.
Теоретические представления Толмена характеризовались трезвой логичностью и одновременно достаточной изощренностью. Они были подкреплены экспериментами, которые часто бросали вызов другим теориям и поэтому трудно с ними уживались. Критическим высказываниям о том, что теории Толмена «похоронят животное в размышлениях» (Guthrie, 1952), не дав ему возможности «предугадать» своего поведения, можно противопоставлять тезис о том, что, согласно молекулярному бихевиоризму, животное должно «погибнуть в деятельности».
Язык и психические представления
Способны ли животные к психической деятельности, подобной психической деятельности человека, или они просто бессмысленные автоматы? На протяжении последних ста лет этот вопрос постоянно занимал умы психологов и этологов, и господствующее мнение «металось» от одной крайности к другой. Наши знания по физиологии и поведению животных неизмеримо выросли, но чем больше мы узнавали, тем более сложной представлялась нам эта проблема. В настоящей главе мы свели воедино некоторые системы аргументов, которые были выработаны в пределах различных зоопсихологических школ, и попытались сформировать такую позицию, с которой любой студент оказался бы в состоянии составить свое собственное мнение по поводу психической деятельности животного.
По-видимому, существует много аспектов в поведении человека, которые отличают его от животного. Ранее считали, что только человек способен создавать и использовать орудия, однако теперь мы знаем, что этой способностью обладают многие виды животных. Создается впечатление, что по мере того как увеличиваются наши знания о поведении животных, различия между человеком и животными начинают сокращаться. Вместе с тем некоторые способности, которыми обладают люди, обнаружить у животных очень трудно. Одна из таких способностей - язык.
Нам кажется, что наличие языка - это уникальное свойство человека. Возможно, что это и не единственная черта, выделяющая нас из мира животных. Так это или не так, но мы не должны позволить нашим антропоморфическим чувствам затуманивать наше представление о возможности существования языка у животных. К сожалению, в течение долгого времени существовала тенденция определять язык таким образом, что создавалась уверенность, будто это исключительная привилегия людей. Самые яркие сторонники этих представлений утверждали, что для языка необходимо сознание, которым обладает только человек, или что язык зависит от речи, к которой опять-таки способен только человек. С научной точки зрения такой подход недопустим, поскольку он вносит в исследование непреодолимые предубеждения.
К сожалению, определить понятие языка с объективной точки зрения очень нелегко, поскольку он характеризуется многими необходимыми признаками. Например, мы можем согласиться с тем, что язык - это средство коммуникации, но что не все средства коммуникации являются языком. Человеческий язык обычно существует в форме речи, но это не всегда так, например в случае с азбукой Морзе. Язык использует символы, но символичны и некоторые аспекты коммуникации у пчел. Язык осваивают в течение специфического чувствительного периода развития, но то же самое наблюдается у некоторых птиц, научающихся песне своего вида. С помощью языка можно передавать информацию не только о сиюминутных ситуациях, но и о таких, которые оказываются удаленными и во времени, и в пространстве. Но некоторые сигналы тревоги у животных обладают теми же свойствами. По-видимому, некоторые аспекты языка, например правила грамматики, выделяют его из других видов поведения животных, но даже и этот тезис достаточно спорный. Когда мы изучаем вопрос о существовании языка в царстве животных, мы должны продвигаться очень осторожно.
26.1. Невербальная коммуникация у человека
Дарвин (1872) понимал, что между выражениями лица человека и мимическими реакциями животных, особенно приматов, существует много общего. В настоящее время ученые широко признают, что напрашивается прямое сравнение между невербальными аспектами человеческой коммуникации и демонстрациями у животных. Однако следует учитывать, что некоторые стороны невербальной коммуникации людей непосредственно связаны с языком. Очевидный пример - знаковый язык, которым пользуются глухонемые. Однако и многие простые жесты вроде поднятого большого пальца руки — жеста - знака, вероятно, берут свое начало непосредственно в языке.
В гл. 22 мы уже видели, что многие демонстрации и мимические реакции животных представляют собой результат ритуализации. В процессе эволюции они, по-видимому, произошли от оборонительных реакций, движений намерения, смещенной активности животных и т. д. Имеются ли у человека какие-то эквивалентные стереотипные демонстрации?
Одна из проблем в исследовании человеческой коммуникации заключается в том, что она может иметь различные особенности, связанные с культурными традициями народа. Например некоторые способы выражения согласия или отрицания чего-либо содержат такие движения головы, как кивание или небольшие повороты головы из стороны в сторону. Однако смысл, который приписывается этим движениям, широко варьирует в зависимости от культурных традиций народа (Leach, 1972). Например, в Греции и некоторых других странах отрицание «нет» выражается сильным и резким движением головы назад. Тем не менее при кросскультуральном обследовании было установлено, что некоторые выражения лица являются универсальными и, по-видимому, имеют одно и то же значение у народов самых различных культур, например «взлет» бровей (рис. 26.1), когда обе брови мгновенно поднимаются вверх. Обычно этот сигнал подается как форма приветствия на расстоянии (Eibl-Eibesfeldt, 1972). У разных народов встречаются очень небольшие различия в использовании этого сигнала. В Европе его используют для приветствия хороших друзей и родных, в Новой Гвинее - для приветствия иностранцев, в Японии эта мимическая реакция подавлена и считается неприличной. В целом же взлет бровей используется как форма дружелюбного приветствия или одобрения, но у сдержанных и подозрительных лиц ее может и не быть (Eibl-Eibesfeldt, 1972).

Рис. 26.1. «Взлет бровей - пример мимической реакции у человека.
Целый ряд основных мимических выражений, таких, как улыбка, смех и крик, - универсален. Они встречаются не только у представителей различных культур, но и у людей, родившихся глухими и слепыми (Eibl-Eibesfeldt, 1970); особенно выражены они у маленьких детей (Blurton-Jones, 1972). Эти мимические реакции человека можно непосредственно сравнить с мимическими реакциями других приматов. При анализе различных типов выражения лица человека было обнаружено, что существует целый ряд условий, в которых проявляются достаточно стереотипные реакции (Hoof, 1972, 1976). К ним относится настороженность, которая выражается в относительно фиксированном пристальном взгляде и некотором напряжении лицевых мышц. Такое выражение лица очень напоминает соответствующую мимическую реакцию других приматов, за исключением того, что у некоторых и уши также могут быть направлены в сторону того, что привлекло их внимание. Чувство удивления внешне проявляется в том, что человек на какое-то время поднимает брови, открывает глаза, а зачастую и рот. Это выражение лица, по-видимому, не имеет никакого аналога среди обезьян. Проявления страха сходны у многих приматов: для них характерны широко раскрытые глаза и сжатые губы. При чувстве отвращения человек морщит нос и поднимает верхнюю губу, прищуривает глаза и отворачивает лицо. Все компоненты этого поведения - производные защитных реакций, направленных на то, чтобы избежать вредных раздражителей. У человека внешнее выражение отвращения ритуализовано, но у других приматов это просто набор защитных реакций, которые в коммуникации не играют никакой особой роли. При печали брови изгибаются дугой, углы рта вытягиваются, опускаются вниз и в стороны, кривя губы. Если печаль велика, могут быть и слезы.
По-видимому, такое выражение печали у человека, а возможно, и у шимпанзе ритуализовано. Со стороны других членов группы оно вызывает реакции утешения. Гнев может проявляться по-разному, но обычно для него характерны прижатые к оскаленным зубам губы, пристальный взгляд и нахмуренное лицо. Радость выражается улыбкой и смехом (рис. 26.2), которые часто ассоциируются с юмором и поэтому считаются исключительной принадлежностью человека. Однако некоторые этологи оспаривают эту точку зрения (например, Hoof, 1972; Eibl-Eibesfeldt, 1970).
У обезьян имеется демонстрация расслабления, при которой открывается рот; она связана с игривостью (рис. 26.3). Внешне эта мимическая реакция похожа на улыбку человека; однако между ними существует и ряд различий, особенно в характере сопровождающего их дыхания. И у обезьян, и у человека эта напоминающая улыбку демонстрация возникает в ответ на что-то неожиданное в окружающей обстановке. У шимпанзе и маленьких детей она имеет чисто физическую природу, тогда как у взрослых людей природа этой демонстрации может быть и физической, и чисто интеллектуальной. Однако в обоих случаях эта мимическая реакция характеризует, как правило, социальные взаимодействия. Улыбка человека - это не просто слабый смех (хотя эти две реакции часто взаимосвязаны); улыбка возникает также и в таких ситуациях, когда дело касается легкого страха или опасения, как, например, в случае социального приветствия и ободрения. У многих приматов беззвучное обнажение зубов - это типичное проявление мимики субординантного животного, означающее признание или подчинение доминантным членам группы. Однако у шимпанзе это ободряющее и дружелюбное выражение сродни человеческой улыбке.

Рис. 26.2. Маленький ребенок, которого щекочут, смеется с открытым ртом. Эта разновидность смеха - с открытым ртом, без оттягивания назад уголков рта - у взрослых становится редкой. (Из The Oxford Companion to Animal Behaviour, 1981.)

Рис. 26.3. Капуцин расслабленно раскрывает рот, когда человек его щекочет. (Из The Oxford Companion to Animal Behaviour, 1981.)
Мимические реакции человека и других млекопитающих обычно рассматривают как выражения эмоции, или аффекта (Ekman, 1971). Однако многие другие типы невербальной коммуникации можно классифицировать и по-другому. Помимо аффективной мимики, можно выделить и другие классы невербального общения (Ekman, Friesen, 1969), в том числе адаптеры, которые выполняют и коммуникативные, и некоммуникативные функции. Примерами могут служить присущие животным движения чистки тела и движения намерения. К классу эмблем относятся невербальные движения, у которых имеются свои вербальные аналоги. Они составляют знаковые языки, которыми пользуются глухонемые. Кроме того, к эмблемам относятся неприличные жесты и различные сигнальные движения, используемые на расстоянии, например подзывающие жесты. Класс иллюстраторов включает движения говорящего, которые иллюстрируют отдельные моменты его речи. К ним относятся жесты выразительные, указующие и пр. Класс регуляторов объединяет жесты, которые используются для регулирования речевого потока у двух разговаривающих друг с другом людей. Примерами могут служить кивание головой, движения, связанные с общением посредством глаз, а также различные изменения позы тела.
Эрджайл (Argyle, 1972) предложил что-то вроде упрощенной классификации средств невербального общения, основанной на результатах поведения. Он выделяет три основные категории:
1. Управление ближайшей социальной обстановкой. Сюда относятся жесты и позы, которые передают соответствующее отношение и эмоциональное состояние человека. Так, например, человек может показывать свое чувство превосходства, неприязни, полового влечения или чего-то подобного, не выражая этих чувств словами.
2. Подкрепление речевой коммуникации. Эта категория объединяет классы регуляторов и иллюстраторов, а также различные жесты, наличие или отсутствие которых может изменить смысл произносимых слов. Было показано, что в этом плане особую важность приобретает зрительный контакт (рис. 26.4).
3. Замещение речевой коммуникации. У представителей самых различных культур существуют знаковые языки; такие языки можно использовать, когда речевое общение затруднено или почему-либо невозможно. Общение между охотниками в тот момент, когда требуется тишина, по-видимому, представляет собой одну из первых ситуаций, в которой оказался полезным язык знаков. Современным эквивалентом этого можно считать систему сигналов, которой пользуются рабочие в обстановке сильного шума. Изощренная знаковая система, в основе которой лежит язык, используется для коммуникации на расстоянии (например, семафор) и при общении глухонемых (например, американский язык знаков, или амеслан, - AMErican Sign LANguage, or Ameslan). Письмо и азбуку Морзе, - по-видимому, следует также отнести к этой категории.
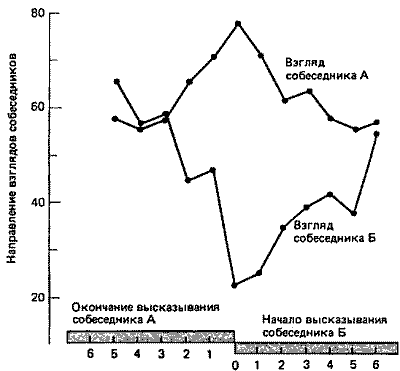
Рис. 26.4. Направление взгляда (ордината) в начале и конце длительного высказывания - это важный аспект невербальнои коммуникации между двумя беседующими людьми. Одинаковое направление взгляда свидетельствует о зрительном контакте (eye contact). Обратите внимание на то, что дальше всего собеседники отводят друг от друга взгляды в тот момент, когда речь одного из них сменяется речью другого. (По Kendon, 1967.)
Можно выделить два основных типа невербального общения. Прежде всего это такое невербальное общение, которое помогает использованию языка или является частью языка. Второй тип коммуникации не зависит от языка; он близок тому виду общения, которым пользуются животные подавляющего большинства видов.
26.2. Язык
В общении животных можно обнаружить ряд особенностей, характерных для языкового общения человека. Например, сигналы, используемые в языке человека, весьма произвольны, так как по своим физическим особенностям они не похожи на те характеристики окружающего мира, которые они обозначают. Это абстрактное качество обнаружено также и в коммуникативном поведении медоносных пчел (Apis melliferd), исследование которого впервые предпринял Карл фон Фриш (см. гл. 23).
Танец медоносной пчелы во многих отношениях является символическим. Скорость виляющего танца указывает на расстояние источника пищи от улья, при этом точное соотношение между скоростью исполнения танца и расстоянием определяется местными «договоренностями». По-видимому, различные географические расы пчел используют различные «диалекты». Так, один и тот же элемент виляющего танца обозначает примерно 75 м у немецкой медоносной пчелы, около 25 м у итальянской пчелы и всего лишь 5 м у пчелы из Египта. Если все пчелы в колонии придерживаются данной договоренности, то не имеет никакого значения, какому именно расстоянию соответствует элемент их танца. Некоторые исследователи (например, Hinde, 1974) считают, что, поскольку используемые пчелами символы (направление и скорость танца) физически связаны с направлением на источник пищи, они не могут считаться произвольными. Однако в любой системе символов, которая представляет какой-то диапазон величин, соответствие между областью символов и областью соответствующей реальности является ограниченным.
Другие этологи (например, Gould, Gould, 1982) рассматривают танец медоносной пчелы как пример произвольного «соглашения», доказывая, в частности, что вместо солнца в качестве точки отсчета пчелы могут использовать направление на север. Танец медоносной пчелы несет информацию о ситуациях, которые имеют место вдали от общающихся животных, - эту особенность многие считают важным свойством. Присущим языку человека. Танец сообщает не только об источниках пищи, удаленных в пространстве (иногда до 10 км), но и о тех, которые пчелы посетили несколько часов назад. В течение всего этого времени пчела-сборщица сохраняет психический образ траектории движения солнца и в соответствии с этим корректирует свой танец.
Другая особенность языка человека состоит в том, что он представляет собой открытую систему, в которую могут включаться новые сообщения. Пчелиный танец может сообщить о новых источниках пищи, но, по-видимому, это пример достаточно ограниченной «открытости». Однако пчелы используют танец также и для того, чтобы направить соплеменницу к воде, прополису (особым древесным выделениям, используемым для замазывания отверстий в улье), а возможно (в период роения), и к новым местам, где можно будет обосновать гнездо (Gould, 1981).
Конечно, некоторые особенности языка человека, например его акустические свойства, отсутствуют в языке пчелы. Но некоторые из этих особенностей можно увидеть в проявлениях коммуникации у других животных, например в песне птиц. Несомненно, что язык человека более сложен и изощрен, чем язык животных. Но означает ли это, что между общением людей и общением животных существует качественная разница, или здесь все дело в степени? На этот вопрос еще нет однозначного ответа.
Если животные способны пользоваться языком, тогда можно ожидать, что ближе всего к людям в этом отношении будут высшие обезьяны. У этих животных голосовые реакции и мимические движения отличаются утонченностью и сложностью. Поэтому можно предположить, что они разговаривают между собой на языке, нам пока непонятном. Были предприняты различные попытки установить, способны ли высшие обезьяны пользоваться языком, которым пользуемся мы. Прежде всего попытались научить шимпанзе копировать человеческую речь. Орангутан после нескольких лет обучения оказался способным произносить только два слова: «папа» и «сир» (чашка). Потратив еще больше времени на тренировку, шимпанзе Вики справилась со словами «папа», «мама», «сир» и «up» (вверх) (Hayes, Nissen, 1971). В обоих случаях обезьяны произносили слова очень нечетко, и стало очевидным, что у этих животных просто нет голосового аппарата, с помощью которого можно было бы воспроизводить звуки человеческой речи. У шимпанзе и плода человека гортань расположена в верхней части голосового пути, тогда как у взрослых людей - в нижней его части (рис. 26.5). Такое расположение гортани и дает возможность человеку изменять с помощью языка конфигурацию полости глотки и таким образом производить широкий спектр модулированных звуков. Шимпанзе и другие человекообразные обезьяны просто не способны производить эти звуки (Jordan, 1971; Lieberman, 1975).
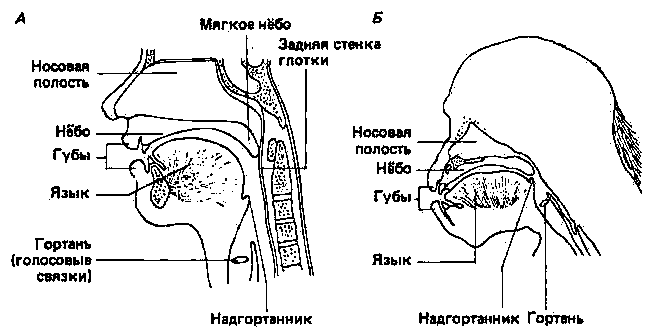
Рис. 26.5. Схематическое изображение (сагиттальное сечение) головы и шеи взрослого человека (А) и взрослого шимпанзе (Б). (Lieberman, 1975.)
И хотя ясно, что человекообразные обезьяны не могут говорить, вес равно в отношении их коммуникации остается много невыясненных вопросов. Есть ли у них для общения что-то вроде языка? Способны ли они понимать звуки человеческой речи? Можно ли их научить пользоваться языком человека с применением каких-то, других средств вместо речи?
Звуковой репертуар шимпанзе насчитывает около 13 звуков, но они могут издавать и звуки с какими-то промежуточными характеристиками. Обезьяны используют эти звуки как для дистанционного общения, так и при близком взаимодействии. Они различают голоса знакомых особей и постоянно используют звуки для поддержания контакта друг с другом, когда находятся в густом подлеске, или при наличии каких-то других препятствий, мешающих видеть своих соплеменников. На основе движений намерения, мимических реакций, запахов и звуков, которые производит какая-то обезьяна, другие животные группы могут ее опознать, определить, где она находится, каково ее мотивационное состояние и, вполне вероятно, чем она занимается. Однако здесь нет никаких признаков истинного языка, и поэтому нам нужно поискать данные о том, что животные могут с помощью символов обмениваться информацией о внешнем мире.
Было обнаружено, что некоторые виды животных издают сигналы тревоги, которые различаются в соответствии с видом опасности. Взрослые зеленые мартышки (Cercopithecus aethiops) производят различные тревожные звуки, когда увидят питона, леопарда или африканского воинственного орла. Другие обезьяны, услышав эти звуки, предпринимают действия, соответствующие характеру обнаруженной опасности. Если это змея, то они начинают смотреть вниз, а если орел, то, напротив, - вверх. Если они слышат сигнал, предупреждающий о близости леопарда, они спасаются бегством в ветвях деревьев (Seyfarth, Cheyney, Marler, 1980). Эти наблюдения свидетельствуют о том, что обезьяны способны обмениваться информацией о внешних стимулах, но мы не можем быть уверены в том, что они не сообщают друг другу всего лишь о различных эмоциональных состояниях, вызванных этими стимулами.
Мензел (Menzel, 1974; 1979) провел с обезьянами шимпанзе эксперименты, в которых выяснил, могут ли эти животные передавать друг другу информацию о местоположении пищи. На шести шимпанзе, содержащихся на определенном участке поля, он провел серию тестов. В сопровождении одной из подопытных обезьян он прятал пищу в поле, а затем выпускал всех шестерых животных, предоставляя им возможность разыскать эту пищу. Как правило, вся группа с восторгом направлялась бегом прямо к пище и очень быстро находила ее. Однако обезьяна, которая была свидетелем прятания пищи, отнюдь не всегда возглавляла эту группу. Когда Мензел вместо пищи спрятал змею, шимпанзе приближались к ней очень осторожно, с явными признаками страха. В одном из экспериментов Мензел показывал два разных тайника с пищей двум обезьянам. Когда всех обезьян выпускали, то они обычно выбирали из этих двух тайников наиболее привлекательный. Предпочтение отдавалось либо большему количеству пищи в этом тайнике по сравнению с другим, либо фруктам по сравнению с менее любимыми овощами.
По-видимому, шимпанзе на основе поведения своих компаньонов могут делать заключения об особенностях окружающего их мира. Обезьяна, которой стало известно, где находится пища, своими действиями и эмоциями показывает другим животным степень желаемости цели и направление к ней. Прямых указаний о местонахождении пищи нет, однако другие шимпанзе оказываются достаточно рассудительными, чтобы сделать свое собственное умозаключение об этом (Menzel, Johnson, 1976). Некоторые данные свидетельствуют о том, что в лабораторных условиях эти обезьяны могут научиться указывать на определенные объекты (Terrace, 1979; Woodruff, Premack, 1979) и использовать указующие жесты экспериментаторов как ключ для определения местонахождения пищи (Menzel, 1979). Однако при общении между собой они, по-видимому, не используют указательных жестов или каких-либо других знаков направления.
26.3. Обучение высших обезьян «разговаривать»
Хотя и кажется очевидным, что шимпанзе обычно не сообщают друг другу об объектах, которые удалены от них во времени и в пространстве, но вполне может быть, что их можно научить этому. Если бы мы знали о том, до какой степени человекообразные обезьяны способны использовать различные особенности языка, мы, вероятно, смогли бы кое-что узнать и о наших собственных способностях.
В семье Келлогов (Kellog, Kellog, 1933) дома жила шимпанзе по имени Гуа. Они утверждали, что она научилась понимать 95 слов и фраз, когда ей было 8 мес, т. е. примерно столько же, сколько их сын Доналд, который был на три месяца старше. Для проверки способностей Гуа ей давали карточку, на которой были изображены 4 картинки. Другая шимпанзе по имени Элли жила в доме с людьми и научилась немного понимать речь. Ее научили некоторым жестам, которые соответствовали определенным словам в языке амеслан. Она была способна, услышав произнесенное слово, делать правильный знак (Fouts et al., 1976). На основе других экспериментов, которые проводились на горилле (Patterson, 1978) и собаке (Warden, Warner, 1928), было высказано предположение, что эти животные способны сформировать связь между звуками и зрительными сигналами.
Некоторые ученые попытались изучить, до какой степени приматы могут произвольно управлять звуками, которые они издают. В одном из опытов макак-резус должен был на включение зеленого света «лаять», а на включение красного - «ворковать». Обезьяны научились правильно производить эти звуки, чтобы получать пищевое подкрепление (Sutton, 1979). Орангутан научился издавать три различных звука, чтобы получать пищу, питье или возможность контактировать с хозяином (Laidler, 1978), а шимпанзе научилась лаять, чтобы побудить человека поиграть с ней (Randolph, Brooks, 1967). В качестве контроля шимпанзе научили начинать игру с женщиной-экспериментатором двумя различными способами: прикасаясь к ней, когда она оказывалась лицом к обезьяне, и издавая воркующие звуки, когда женщина стояла к обезьяне спиной. На основе этих экспериментов пришли к заключению, что обезьяны в определенных пределах способны осуществлять произвольное управление над издаваемыми ими звуками. Если с ними не проводить специальной тренировки, то они, скорее всего, не будут подражать звукам, которые слышат, даже в том случае, когда живут вместе с людьми, в семейной обстановке (Kellog, 1968). Однако тот факт, что они с готовностью подражают действиям человека, наводит на мысль, что звуки - это не то средство, которое готовы были бы освоить человекообразные обезьяны, чтобы использовать его для общения, выходящего за пределы их обычного ограниченного репертуара (Passingham, 1982).
Как только стало ясно, что речь не является необходимой составляющей языка и что способность издавать звуки или отвечать на них не обязательно связана с ответом на вопрос: «Способно ли животное овладеть языком?» - сразу открылся путь для исследования языка с помощью манипулирования зрительными символами (Gardner, Gardner, 1969; Premack, 1970). Гарднеры начали работать с одной обезьяной шимпанзе по имени Уошо; они обучили ее языку амеслану, в котором слова представлены в виде жестов пальцев и руки. Уошо обучалась с 11 мес до 5 лет и за это время освоила 132 знака (Gardner, Gardner, 1975). Уошо самостоятельно научилась комбинировать эти знаки в цепочки из 2-5 слов. Первыми такими комбинациями были «открой» и «дай сладкого» (Gardner, Gardner, 1971). После этого у Гарднеров были еще две обезьяны шимпанзе. Они обучались с самого рождения и делали это гораздо быстрее, чем Уошо. Фаутс (Fouts, 1975) также обучал шимпанзе. А Паттерсон (Patterson, 1978) научил одну гориллу использовать знаки, производимые кистью руки, и отвечать на голосовые команды, которые подавались на английском языке. Террейс (Terrace, 1979) со своими сотрудниками научил шимпанзе по имени Ним Чимпски пользоваться языком амеслан; в течение всего эксперимента они тщательно расшифровывали каждый знак, который делал Ним, и каждую комбинацию знаков.
Вопрос: "А (бутылка) — это то же самое, что Б (карандаш) ?"
Варианты ответов: "Да" или "нет"
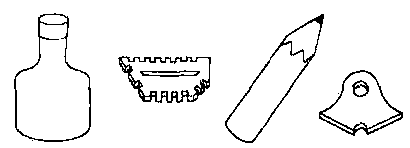
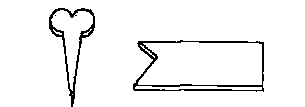
Рис. 26.6. Пример вопроса, который задавали шимпанзе Саре посредством пластмассовых жетонов. Сара могла отвечать, выбирая один из изображенных внизу вариантов ответа. (По Premack, 1976.)
Примак (Premack, 1976, 1978) научил шимпанзе по имени Сара читать и писать. Для этого он использовал цветные пластмассовые жетоны различной формы, которые символизировали слова. По своей конфигурации эти жетоны никак не напоминали те вещи, которые они символизировали (рис. 26.6). Они располагались на вертикальной магнитной доске, и Сара могла отвечать на вопросы, помещая на эту доску соответствующие фигуры. Сара освоила 120 пластмассовых символов, хотя ее никто не заставлял осваивать столь обширный словарь (Premack, 1976). Она могла выполнять команды и отвечать на вопросы, используя комбинации из нескольких символов. По этой методике Примак и его коллеги обучали и других шимпанзе.
Дьюэйн Румбо (Duane Rumbaugh, 1977) применяла другой метод обучения. Она использовала искусственную грамматику, названную «йеркиш» (Jerkish) (Glasersfeld, 1977). Шимпанзе по имени Лана научилась оперировать клавиатурой компьютера, с помощью которой на экран выводились символы слов. Компьютер был запрограммирован таким образом, чтобы распознавать, соответствует ли правилам грамматики использование этих символов или они употребляются неправильно; в соответствии с этим Лана получала подкрепление. Преимущество этого подхода к обучению заключалось в том, что Лана имела возможность общаться с компьютером в любое время дня, а не ожидать установленных часов эксперимента. И другие шимпанзе также научались общаться друг с другом с помощью метода, основанного на использовании компьютера (Savage-Rumbaugh et al., 1978; 1980).
Оценивая результаты этих экспериментов, необходимо иметь в виду и тот факт, что шимпанзе могут обманывать. Они способны использовать непроизвольные намеки, которые могут допускать экспериментаторы, или могут просто научиться последовательности трюков подобно тому, как это делают животные в цирке.
В 1978 г. Гарднеры проводили эксперименты с Уошо в таких условиях, когда сами экспериментаторы не знали ответа на вопрос, предлагаемый обезьяне. Уошо должна была назвать объект, показываемый на слайде, делая соответствующий знак находящемуся рядом человеку, который не мог видеть этого слайда. Второй экспериментатор мог видеть жесты Уошо, тогда как сама Уошо его не видела; при этом экспериментатор не видел слайдов. Уошо должна была назвать 32 предмета, каждый из которых ей показывали четыре раза. Она дала правильные ответы на 92 из 128 вопросов. Подобные тесты проводились и на некоторых других шимпанзе, которые были объектами этих исследований (Rumbaugh, 1977; Premack, 1976; Patterson, 1979).
Вполне возможно, что шимпанзе научаются тому, что они должны делать при получении определенных сигналов, точно так же, как цирковые животные обучаются тому, что им следует делать в ответ на соответствующие сигналы дрессировщика. Чтобы определить, понимают ли шимпанзе смысл знаков и символов, которыми они манипулируют, необходимо провести такой эксперимент, где обезьяне пришлось бы называть предметы в ситуации, отличающейся от той, где происходило научение. В этом плане было проведено множество различных опытов (например, Gardner, Gardner, 1978; Savage-Rumbaugh et al., 1980), результаты которых показали, что шимпанзе на самом деле способны называть предъявляемые предметы. Более того, иногда обезьяны делали это спонтанно. Так, например. Ним делал знак собаки, когда видел живую собаку или ее изображение или когда слышал собачий лай (Terrace, 1979).
Существуют определенные доказательства того, что шимпанзе могут постигать смысл слов, т.е. что они на самом деле способны употреблять названия различных объектов. Однако их способности не столь очевидны, когда дело касается многих других аспектов языка человека, которые представляют интерес прежде всего при оценке когнитивных способностей шимпанзе. Особый интерес представляет вопрос о том, способны ли шимпанзе вводить в свой репертуар новые (не выученные ранее) «сообщения». Этот вопрос представляется важным и для оценки танца пчел (см. гл. 23).
По-видимому, иногда шимпанзе создают новые фразы. Сообщалось, в частности, о том, что Уошо выдумала слово "candy drink" («сладкое питье») для обозначения арбуза, а лебедя назвала "water bird", т. е. водяной птицей. Однако такие случаи трудно интерпретировать, поскольку существует возможность, что кажущееся новым использование слова является лишь результатом простой генерализации. Например, Уошо научили знаку цветка, когда показывали ей настоящий цветок. Она освоила этот знак, но пользовалась им не только в отношении цветка, но и в отношении аромата табака и запахов кухни. Возможно, что Уошо связала этот знак с запахом цветка и генерализовала его на другие запахи (Gardner, Gardner, 1969).
Другая проблема состоит в том, что шимпанзе иногда создают новые комбинации слов, которые выглядят как не имеющие никакого смысла. Любимой пищей Нима были бананы, и он часто комбинировал это слово с другими словами, такими, как питье, щекотание и зубная щетка. Хотя и можно предположить, что "banana toothbrush" («банан» - «зубная щетка») - это требование банана и зубной щетки, чтобы почистить зубы после съедания банана, но это предположение кажется маловероятным, поскольку банан и зубная щетка никогда не оказывались в поле зрения обезьяны в одно и то же время и Ним никогда не просил тех предметов, которых он не мог видеть раньше (Ristau, Robbins, 1982). По-видимому, такие причудливые комбинации слов представляют собой пример игры словами, которая напоминает подобную игру у детей. Экспериментаторы заметили, что Уошо делала знаки и сама себе, когда играла одна, - почти так же, как дети разговаривают сами с собой.
Таким образом, мы можем сказать, что попытки научить шимпанзе и других человекообразных обезьян различным типам человеческого языка имели ограниченный успех. Вероятно, человекообразные обезьяны способны достичь в этом лишь уровня маленького ребенка. Вполне возможно, что различие между человекообразными обезьянами и человеком - это всего лишь различие в интеллекте. Однако вполне вероятной представляется и гипотеза о том, что люди обладают каким-то врожденным аппаратом для освоения языка. Эту мысль первым высказал Хомски (Chomsky, 1972). Во всяком случае, описанные здесь эксперименты с человекообразными обезьянами определенно открыли нам такие их способности, о которых мы раньше и не подозревали, и существенно приблизили нас к пониманию когнитивных возможностей этих животных.
26.4. Истоки человеческого языка
Скорее всего, истоки человеческого языка навсегда останутся для нас загадкой. Брешь, разделяющая лингвистические возможности человека и животных, настолько велика, что мы можем узнать лишь крупицу, когда проводим сравнительные исследования с различными живущими ныне видами животных. Изучение ископаемых остатков дает нам какую-то путеводную нить, но эти данные очень трудно интерпретировать. Проблема заключается в том, что нам не ясно, что именно считать доказательством языковой способности (Passingham, 1982). Если считать, что речь была необходимым предшественником языка, то следует пользоваться данными, относящимися к эволюции голосового аппарата. Реконструкции, основанные на характеристиках костей черепа и нижней челюсти, дают основание предположить, что голосовые аппараты неандертальца, австралопитеков и шимпанзе очень схожи (Lieberman, 1975). Даже если мы не примем в расчет сомнения в надежности таких данных, мы не сможем сделать вывод, что неандерталец не был способен к языку.
Вполне возможно, что эволюция языка началась с какого-то изощренного использования жестов, а речь возникла уже позже (Passingham, 1982).
Слепки мозга ископаемых гоминид дают нам информацию о размерах и форме мозга, которые можно сравнить с соответствующими характеристиками мозга современного человека. Можно предположить, что крупный мозг, вероятнее всего, ассоциируется с языковыми способностями у приматов, однако неизвестно, насколько большим для этого должен быть мозг. Мозг неандертальца слегка превышает по размеру средний мозг человека, но означает ли это, что язык был характерной чертой, присущей жизни неандертальца? Большой мозг может быть признаком интеллекта, однако может оказаться, что «обладание человеческим языком связано с особым типом психической организации, а не просто с большей степенью интеллекта» (Chomsky, 1972). Однако при анализе конфигурации мозга можно получить дополнительные ключи к пониманию этой проблемы. У некоторых гоминид наблюдается асимметрия полушарий - особенность, сходная с особенностью мозга современного человека (LeMay, 1976). У Homo sapiens прослеживается определенная связь между асимметрией мозга и существованием языка, и вполне возможно, что нечто подобное имело место у некоторых ранних гоминид (Passingham, 1982). Были также предприняты попытки идентифицировать в слепках мозга ископаемых некоторые структуры, относительно которых известно, что у современного человека они имеют отношение к языку (Holloway, 1976). Быть может, с помощью более совершенных методик (например, Holloway, 1981) можно было бы более глубоко провести такого рода анализ, однако те данные, которыми мы располагаем в настоящее время, подтверждают только самые осторожные предположения.
Различные предметы, изготовленные древним человеком, которые обнаружены на местах его стоянок, свидетельствуют об использовании им символических изображений. Каменные орудия и другие инструменты уходят в глубь веков на 2,5 млн. лет (Lewin, 1981), но они еще не являются необходимыми свидетелями существования языка (Passingham, 1982). Украшения, обнаруживаемые на ископаемых изделиях, заставляют предположить, что их изготовитель владел символическим языком (Marshack, 1976), и такие украшенные изделия отстоят от нас примерно на 300 000 лет. Однако рисунки-изображения, по-видимому, появились только у Homo sapiens около 30 000 лет назад. Письменный язык, скорее всего, развивался от изобразительных символов с эмоциональным и мистическим значением к более информативному изображению каких-то конкретных ситуаций (рис. 26.7). Однако мы можем идентифицировать свойства истинного языка только тогда, когда обнаружим символы для звуков речи, а не для понятий (КИх, 1982).
26.7. Изображение вражды племен, сделанное индейцами племени Сью. (По Klix, 1982.)
26.5. Язык и познание
Как мы уже видели, шимпанзе и других человекообразных обезьян можно научить разговаривать с человеком, используя язык знаков, или посредством чтения и письма с помощью пластиковых жетонов, или с помощью символов, которые обеспечивает компьютер. Эти данные заставляют предположить, что шимпанзе можно научить значению слов в том смысле, что эти обезьяны смогут использовать названия предметов. Они в состоянии создать словарь более чем на сто слов и, по-видимому, способны иногда составлять новые фразы.
Чрезвычайно интересен вопрос, имеющий множество различных аспектов и заключающийся в следующем: обнаруживаются ли в экспериментах с различными вариантами языка обезьян какие-либо когнитивные способности этих животных. Один из аспектов этого вопроса - различие между знанием «как» и знанием «что». Обезьяна может знать, как выпросить подкрепление в том смысле, что она может научиться делать соответствующий жест. Однако это умение нельзя приравнять к знанию того, что, произведя какой-то определенный жест, можно получить за него награду. «Знать что» ("know that") подразумевает понимание взаимоотношений между явлениями, выходящими за рамки простой связи-стимула и ответа. Человеческое «знание как» ("knowing how") распространяется на очень многие примеры сложного мастерства, такие, как быстрое печатание на машинке или игра в гольф, где исполнитель может добиваться хорошего результата, не понимая или будучи не в состоянии описать связь между целью поведения и выполняемыми действиями. В других случаях, напротив, люди отчетливо понимают весь процесс, который приводит их к определенной цели, и могут его описать. В своих обзорах Ристау и Роббинс (Ristau, Robbins, 1981; 1982) обсуждают вопрос о том, можем ли мы на основе экспериментов с языком понять, каким путем из этих двух действуют человекообразные обезьяны.
Здесь уместно рассказать о результатах некоторых экспериментов, проведенных с Ланой (Rumbaugh, Gill, 1977). Лана научилась пользоваться фразами, составленными из пластмассовых символов, чтобы выпрашивать определенные виды подкрепления. Она могла написать: «Пожалуйста машина дать банан» ("Please machine give banana"), что легко можно интерпретировать как знание того, каким путем можно получить банан. Однако Лану попытались также научить называть два предмета: банан и конфеты фирмы «М и М» (М&М candy). Ей по очереди показывали банан и конфету «М и М» и спрашивали (с помощью компьютерной клавиатуры экспериментатора): «Это называться как?» Правильный ответ (с помощью клавиатуры) был бы: «Это называться банан». Правильные ответы подкреплялись. Лане потребовалось 1600 тестов (сочетаний), чтобы научиться решать эту задачу, несмотря на то что раньше она сотни раз выпрашивала эти лакомства, применяя стандартную фразу: «Пожалуйста машина дать банан». На основе этого возникает предположение, что Лана не знала значения символов, с которыми она раньше манипулировала, выпрашивая конфеты или банан. Однако стоит отметить, что от Ланы первоначально и не требовалось знать значения символов и что стандартное выпрашивание указанных лакомств на основе «знания как» сделалось привычкой, которую трудно изменить. Такую интерпретацию подтверждают данные о том, что при последующей тренировке Ланы, нацеленной на обучение ее называть другие предметы, успеха добились гораздо быстрее. Вполне возможно, что начальные трудности в приобретении Ланой умения называть предметы возникли потому, что она не смогла отличить требования новой ситуации от тех, где символы банана и конфет «М и М» использовались первоначально.
В экспериментах с Сарой Примак (Premack, 1976) следующим образом обучал ее понятию «называется». Перед настоящим яблоком на некотором расстоянии клали его пластмассовый символ. От Сары требовалось, чтобы она заполнила промежуток между ними еще одним пластмассовым жетоном, который должен был обозначать «называется». Таким образом, она явно создавала предложение: «Яблоко (объект) называется яблоко». Понятие «не называется» формировалось за счет присоединения обычного символа отрицания к пластмассовому символу «называется». Когда символ яблока клали на некотором расстоянии перед бананом, от Сары требовалось, чтобы она выбрала правильный символ и заполнила им промежуток между предметами (рис. 26.8). В этом случае Сара получала награду, если выбирала символ «не называется». Она оказалась способной правильно использовать эти символы и в тестах с названиями реальных предметов, и в последовательностях с другими пластмассовыми символами.
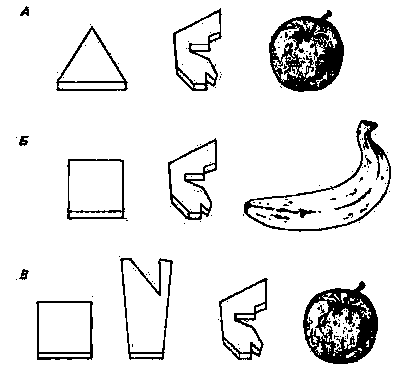
Рис. 26.8. Методика обучения Сары понятию «называется». А. Символ «называется» помещают между символом яблока и настоящим яблоком. Б. Символ «называется» помещают между символом банана и настоящим бананом. В. Между символом банана и яблоком помещают символ «называется», а перед ним - символ «нет». (По Premack, Premack, 1972.)
Способность научаться тому, что абстрактные фигуры являются символами объектов реального мира, предполагает, что у шимпанзе есть какое-то понятие типа «знания что», подобное декларативной репрезентации (declarative representation). Однако трудно придумать какое-то доказательство этого, которое никак нельзя было бы связать с особенностями применяемой методики. Ведь вполне может быть, что Сара обучилась просто тому, что жетоны «называется» и «не называется» означают ответные действия, которые нужно выполнить в тех случаях, когда две вещи (например, яблоко и символизирующий его жетон) будут эквивалентны или неэквивалентны. А это разновидность «методики соответствия» (см. гл. 27.1).
26.6. Психические образы
Вопрос о психическом представлении - это вопрос, который вот уже более десяти лет занимает важное место в когнитивной психологии человека. Методика, которая обычно используется для получения объективной меры такого непостижимого явления, как психические представления, - это психическая хронометрия. Она учитывает время, необходимое для решения какой-то пространственной задачи, как показатель активности соответствующих психических процессов (Posner, 1978). Например, в одном из классических исследований (Shepard, Metzler, 1971) испытуемые должны были рассматривать пары изображений трехмерных объектов. В каждой пробе от испытуемого требовалось указать, одинаковы ли два объекта по форме или один является зеркальным отображением другого. Два объекта могли различаться также и по своей ориентации (рис. 26.9). Было показано, что время, необходимое для того, чтобы определить, одинаковы ли объекты по форме, закономерно возрастало при увеличении угловых различий между парами рисунков, предъявляемых испытуемому (рис. 26.10). Из этих наблюдений обычно делают вывод, что, прежде чем сравнивать по форме оба изображения, испытуемые поворачивают «в уме» мысленный образ одного объекта, чтобы поставить его в одинаковое положение с другим. Хотя истинная природа образа, используемого в этом эксперименте, еще остается спорной (Kosslyn, 1981; Cooper, 1982), наиболее распространенная прямолинейная интерпретация этих данных состоит в том, что здесь имеет место какая-то разновидность психического образа и что процессы вращения и сравнения выполняются один за другим. Тот факт, что время реакции является функцией от углового смещения предъявляемых изображений, служит основанием для заключения о том, что мысленное вращение объекта занимает определенное время (около 30 мс на каждые 20 градусов) (Cooper, Shepard, 1973).
Проверка способности к зрительному распознаванию объектов независимо от их относительной пространственной ориентации используется в нескольких тестах для оценки интеллекта и способностей личности (Petrusic et al., 1978); было бы интересно сравнить на этой основе представителей разных видов животных (см. разд. 27.1). Однако до сих пор систематические исследования проведены на животных только одного вида, и результаты представляются достаточно удивительными.
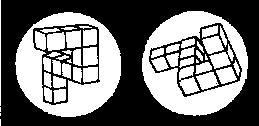
Рис. 26.9. Пример пары фигур, которые используются для «психологической хронометрии». Это идентичные фигуры, но как бы повернутые на 80° одна относительно другой. (Shepard, Metzler, 1971.)
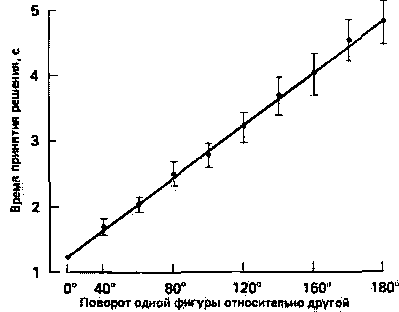
Рис. 26.10. Время, которое требуется для установления идентичности двух схожих фигур, как функция ориентации этих фигур по отношению друг к другу. (Shepard, Metzler, 1971.)
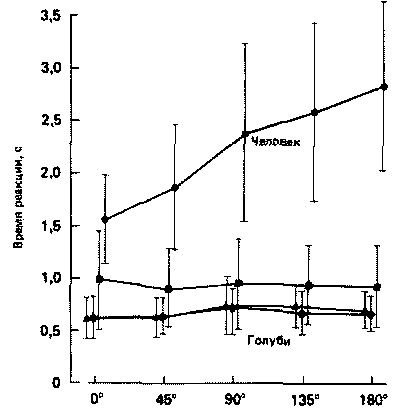
Валери Холлард и Джуан Делиус (Valerie Hollard, Juan Delius, 1983) обучали голубей в камере Скиннера различать прямое и зеркальное изображения фигур, которые предъявлялись птицам ориентированными различным образом (см. рис. 26.11). Затем они измеряли время реакции голубя в тестах на инвариантность по отношению к повороту. Когда эта часть исследований была завершена, камеру размонтировали и тестовую панель, на которой находились зрительные сигналы и клавиши, использовали в аналогичных экспериментах на людях. Таким образом, на основе одних и тех же конфигураций стимулов можно было непосредственно сравнить, как справляются с этой задачей голубь и человек.

Рис. 26.11. Экспериментальная установка (вверху), с помощью которой голубям демонстрировали различные символические изображения (внизу). Птицы были обучены указывать, какая из двух предъявляемых для сравнения фигур больше всего похода на образец. Предъявляемые для сравнения фигуры были повернуты соответственно на 0, 45 и 180°. (Hollard, Delius, 1983.)
Результаты показали, что если судить по сделанным ошибкам, то люди и голуби способны решать эти задачи примерно одинаково хорошо. В то же время были выявлены и определенные различия. У человека время реакции возрастает по мере роста углового несоответствия между сравниваемыми фигурами, как это уже было показано в исследованиях Купера и Шепарда. А у голубей время реакции оказалось независимым от угла поворота фигур, как это видно на рис. 26.12. По-видимому, голуби способны решать такого типа задачу более эффективно, чем люди, осуществляя, вероятно, какую-то форму параллельной обработки информации. Эти результаты не только приложимы к оценке интеллектуальных способностей животных, они также поднимают вопрос об обоснованности исследований психических репрезентаций. Дают ли опыты с голубями основание для предположения о том, что психические образы (представления) необязательно используются при решении такого рода задачи? Если голуби могут решать такие задачи без использования психических образов, можно ли быть уверенными, что психические образы действительно используются людьми? Или же мы просто считаем, что эти результаты легче объяснить с точки зрения психического представления (см. также Dennett, 1978)?
Угол поворота предъявляемых для сравнения фигур.
Рис. 26.12. Среднее время реакции испытуемого как функция угла поворота предъявляемых для сравнения фигур (по результатам исследования 9 голубей и 22 человек). Обратите внимание на то, что у человека время реакции возрастает при увеличении угла поворота предъявляемых фигур, а у голубей - нет. (Hollard, Delius, 1983.)
26.7. Намеренность поведения
О поведении человека можно сказать, что оно намеренное в том случае, когда оно связано с каким-то представлением о цели, которая направляет поведение. Так, если у меня есть психическое представление о желаемом расположении книг на полке и если это психическое представление направляет мое поведение при расстановке книг, то можно сказать, что я ставлю их намеренно. Если же, однако, я ставлю книги на полку в случайном порядке или просто руководствуясь привычкой, то в этом случае процесс расстановки книг не может быть намеренным. Для того чтобы мое поведение было намеренным, совсем не обязательно, чтобы психическое представление о расположении книг было мною осознано. Хотя категории сознания и намеренности иногда бывают связанными, лучше разбирать их отдельно (Dennett, 1978).
Этологи уже давно выделяют у животных движения намерения (см. разд. 22.1) как показатели того, что именно животное собирается делать. И человек-наблюдатель, и другие особи данного вида могут предугадать будущее поведение животного по его движениям намерения, и, вероятно, неумно полагать, что животные не в состоянии прогнозировать ближайшие шаги в своем собственном поведении (Griffin, 1976). Однако более вероятно, что движения намерения - это просто начальные стадии комплексов поведения, которые остаются незавершенными либо потому, что животное находится в состоянии мотивационного конфликта, либо потому, что его внимание переключается на другие возможные виды поведения. Действительно, трудно представить себе, каким образом животное, обладающее сложным репертуаром активностей, могло бы избежать проявлений таких начальных фрагментов поведения. Хотя некоторые так называемые движения намерения и могли стать ритуализованными в процессе эволюции, это отнюдь не означает, что они являются намеренными в обычном смысле этого слова.
Другая форма поведения животных, которая производит впечатление намеренности, - это наблюдаемая у некоторых птиц отвлекающая демонстрация, демонстрация ложного повреждения организма. Если сидящую на яйцах птицу, например песочника (Ereunetes maurf), потревожит какой-либо наземный хищник, то она может покинуть гнездо и вести себя таким образом, будто она ранена, т. е. волочить якобы сломанное крыло, заманивая подальше от гнезда. Когда птица отведет хищника на безопасное расстояние от гнезда, она внезапно вновь возвращается к своему нормальному поведению и улетает прочь (Brown, 1963; Skutch, 1976). Большинство этологов склонны объяснить этот тип поведения в понятиях ритуализованной демонстрации, однако некоторые исследователи (например. Griffin, 1981) не исключают возможности того, что птицы намеренно ведут себя таким образом. Как отмечает Гриффин, наличие у животных психических состояний или намеренного поведения часто особенно рьяно отрицают в тех случаях, где имеется меньше всего доказательных данных. Трудность заключается в том, какие именно данные считать доказательными. С точки зрения Гриффина, такие данные, вероятнее всего, должны быть получены при исследовании коммуникации у животных, в частности у приматов.
Проясняют ли этот вопрос различные исследования коммуникации у шимпанзе? Вудраф и Примак (Woodruff, Premack, 1979) изучали способность шимпанзе к намеренной коммуникации, создавая ситуации, в которых человек и шимпанзе могли кооперироваться или конкурировать при добывании пищи. Они сообщали друг другу посредством невербальных сигналов о том, где находится спрятанная пища. Когда человек помогал шимпанзе, отдавая ей всю найденную пищу, обезьяна успешно посылала и получала поведенческие сигналы о местонахождении пищи. Когда же человек и обезьяна конкурировали друг с другом и человек забирал себе найденную пищу, шимпанзе научилась вводить в заблуждение своего конкурента, не подавая ему нужных сигналов и не принимая в расчет подаваемые человеком поведенческие знаки, которыми он пытался сбить ее с толку. Такое поведение шимпанзе заставляет предположить, что они способны разгадать цели или намерения человека по его поведению и что у них есть определенные знания о том, как человек воспринимает их собственное поведение.
Каммер (Kummer, 1982) приводит обзор подобного рода очевидных способностей, проявляющихся у свободно живущих приматов, а Гриффин (Griffin, 1981) комментирует случаи (Ruppell, 1969) конкуренции за пищу между матерью и несколькими ее почти взрослыми детьми в семье песца. Так, чтобы первому получить пищу, молодой песец может помочиться прямо на морду своей матери. Испытав несколько таких проделок, мать издает сигнал ложной тревоги и, когда молодняк разбегается, хватает пищу. Гриффин отмечает, что трудно интерпретировать такое поведение, если не допустить существования как у матери, так и у молодняка кратковременных намерений и планов. Однако мы должны задать себе вопрос, действительно ли эти животные принимают в расчет мотивы других особей или они просто обучаются эффективным средствам получения пищи в различных ситуациях.
В случае шимпанзе некоторые данные заставляют предположить, что эти обезьяны действительно способны обманывать. Одним из примеров могут служить наблюдения за развитием жеста указывания - нового поведения, которое обезьяны успешно используют, для того чтобы указать на контейнер с пищей или, обманывая экспериментатора, на пустой контейнер. В лабораторных или полевых условиях шимпанзе редко указывают на что-либо, хотя они прекрасно понимают указующие жесты человека. Второе наблюдение касается шимпанзе Сэди, которая указывала на пустой контейнер, когда у нее спрашивали, где находится пища. Когда человек поднимал контейнер и не обнаруживал там пищи, голова Сэди мгновенно направлялась в сторону другого контейнера, в котором, как она знала, находится пища (Premack, Woodruff, 1978).
Временами создается впечатление, что полемика между бихевиористами и исследователями поведения с точки зрения когнитивной психологии никогда не кончится (см., например, отклики на статью Примака и Вудрафа в журн. «Brain and behavioral sciences», 1978). Похоже на то, что по поводу любой совокупности данных по исследованию поведения каждая сторона выдвигает альтернативные объяснения. Однако Деннетт (Dennett, 1978) отмечает, что поведение сложной системы мы можем рассматривать с нескольких точек зрения, которые не обязательно должны быть противоречивыми. Один из возможных подходов - рассмотрение системы с точки зрения ее «задуманного конструирования» (design stance). Если точно известно, каким образом сконструирована данная система, то можно предугадать, каков будет ее запрограммированный ответ на любую конкретную ситуацию. Говоря обычным языком, мы используем этот подход, когда прогнозируем результат наших манипуляций каким-то объектом с известной функцией, к примеру: «Зажги спичку - и будет свет». В биологии такой подход с точки зрения «предназначения» мы используем при прогнозировании, основанном на теории естественного отбора. Фактически в нашем распоряжении есть два подхода: с позиций максимизации совокупной приспособленности и с точки зрения эгоистичного гена, предложенной Доукинсом (Dawkins, 1976; 1982) (см. гл. 6). Существует еще физический подход, при котором прогнозирование происходит на основе оценки физического состояния системы. В науках о поведении обычно используются два альтернативных подхода: физиологический и с позиции теории систем управления. У каждого из них есть свои преимущества и недостатки, и они скорее дополняют друг друга, чем противоречат один другому (McFariand, 1971).
Деннетт (Dennett, 1978) также предложил подходить к рассмотрению поведения сложной системы с точки зрения его намеренности. Согласно этому подходу, считается, что исследуемая система обладает намерениями и определенной информацией, и ее направляют определенные цели. Предлагая этот подход, Деннетт не пытается отбросить поведенческие или физиологические объяснения, а предлагает более сложный высокий уровень объяснения для поведения систем столь сложных, что другие подходы оказываются непригодными: «Намеренная система - это такая система, поведение которой можно (по крайней мере иногда) объяснить и предсказать, полагаясь на приписываемые ей предположения и желания (и другие черты, имеющие оттенок намерения), т. е. на то, что я буду здесь называть намерениями, включая сюда надежду, страх, собственно намерение, восприятие, ожидание и т. д. По-видимому, в каждом отдельном случае возможны другие пути для предсказания и объяснения поведения намеренной системы, например механической или физической, но подход с точки зрения намерений может быть самым удобным или самым эффективным или, во всяком случае, успешным подходом, который применим к объекту, являющемуся намеренной системой» (Dennett, 1979).
Деннетт (Dennett, 1983) утверждает, что этологи и другие исследователи, изучающие поведение животных с когнитивных позиций, нуждаются в описательном языке и в таком методе, который допускал бы разные трактовки и поддавался бы эмпирической проверке. Он полагал, что теория «систем с намерениями» может выполнить эту роль. Деннетт приводит следующие примеры из работы Сейфарта и его коллег (Seyfarth et al„ 1980): «Зеленые мартышки при обнаружении различных хищников издают различные сигналы тревоги. Если эти крики, записанные на магнитную ленту, проиграть в отсутствие хищников, обезьяны будут вести себя в соответствии с этими сигналами. На сигнал о леопарде они будут прятаться на деревьях, на сигнал о змее смотреть вниз. Взрослые обезьяны издают тревожные сигналы в основном при обнаружении леопарда, африканского воинственного орла и питона, а детеныши сигнализируют криком о «леопарде», заметив любое млекопитающее, об «орле» - при виде многих птиц, а о «змее», сталкиваясь с различными объектами, напоминающими змей. С возрастом и опытом обезьяны начинают лучше классифицировать хищников».
Приняв в качестве исходной позиции представление о наличии намерений у зеленых мартышек, мы рассматриваем животное как «систему с намерениями» и считаем, что у него имеются и предположения, и желания, и разумность - те качества, которые присущи системам соответствующего типа или порядка. Рассматривая поведение обезьяны по кличке Том, которая в присутствии другой обезьяны издает тревожный крик, характерный для обнаружения леопарда, мы можем следующим образом описать его как поведение системы нулевого порядка (т.е. не имеющей намерений). Том склонен к трем типам тревоги: боязни леопарда, орла и змеи, каждая из которых вызывает характерные звуковые реакции, которые издаются животным автоматически, без учета их влияния на других обезьян.
При описании намерений первого порядка можно предположить, что Том хочет заставить другую обезьяну, Сэма, залезть на дерево. Том использует особый голосовой сигнал, чтобы вызвать эту реакцию у Сэма. При описании намерений второго порядка мы делаем следующий шаг и утверждаем, что Том хочет, чтобы Сэм поверил, что поблизости находится леопард и поэтому он должен залезть на дерево. При описании намерений третьего порядка можно утверждать, что Том хочет, чтобы Сэм поверил в то, что Том хочет, чтобы Сэм залез на дерево.
Деннетт утверждает, что вопрос о том, какой порядок намерения считать подходящим, - вопрос эмпирический. Например, представим себе, что одинокий самец зеленой мартышки бродит между группами этих животных и, не слыша других обезьян, молча прячется на деревьях, когда видит леопарда. В этой ситуации мы будем вынуждены отвергнуть описание поведения обезьяны как системы с нулевым уровнем намерений. Зеленые мартышки могут распознавать голоса каждого из членов своей группы, поэтому если через громкоговоритель воспроизвести записанный на магнитофоне тревожный крик Тома о близости леопарда в такой ситуации, когда Сэм может видеть Тома, тогда у нас могли бы появиться основания отказаться от объяснения поведения с точки зрения намерения третьего порядка. Если бы мы увидели, что Сэм в этих обстоятельствах действительно влез на дерево, то такое поведение было бы нерациональным, а рациональность - это, как мы предполагаем, свойство «системы с намерением» (Dennett, 1983).
Предположение, что такие системы рациональны, дает нам возможность разработать исследовательский прием, который Деннетт (Dennett, 1983) называет «методом Шерлока Холмса».
В рассказе «Скандал в Богемии» противник Шерлока Холмса спрятал очень важную фотографию в комнате, и Холмс пытался узнать, где она находится. Холмс попросил Уотсона бросить в комнату дымовую шашку и громко крикнуть: «Пожар!» Противник Холмса в это время находился в комнате по соседству с той, где Холмс вел наблюдения. Как и следовало ожидать, на этот крик противник бросился в комнату и вытащил фотографию из того места, где она была спрятана. Не каждый мог бы разработать столь остроумный план, позволяющий управлять поведением своего противника, но когда вся эта ситуация уже описана, то кажется очень легким предугадать действия этого противника (Cherniak, 1981).
Однако, как будет видно из следующего рассказа, метод Шерлока Холмса тоже не гарантирует от ошибок. Будучи аспирантом, я проводил опыты на голубях, вызывая у них фрустрацию либо тем, что давал им пищу нерегулярно (иногда давал, иногда нет), либо тем, что показывал им пищу, которую они не могли достать. Один из этих голубей обычно вел себя по отношению ко мне довольно агрессивно, и у меня сложилось впечатление, что ему не нравится, как я с ним обращаюсь. Однажды я невольно провел на нем эксперимент Шерлока Холмса: я позволил этому голубю вылететь из клетки. Он мгновенно подлетел к сплетению электрических проводов, обеспечивающих работу аппаратуры, и начал выдергивать их клювом. Почувствовав легкое потрясение и чувство вины, я вышел из комнаты, чтобы выпить кофе. Вернувшись обратно, я понял по поведению голубя, что он воспринял провода как гнездовой материал и что его агрессивное отношение ко мне было связано с ранними стадиями полового поведения.
Тем не менее такой подход в некоторых обстоятельствах может оказаться полезным. Например, в опытах Примака и Вудрафа (Premack, Woodruff, 1978) Сэди указывала на ящик с пищей, когда в комнату входил человек, помогающий ей получать пищу, и указывала на пустой ящик, когда входил человек, который не делился с ней пищей. Каким образом мы смогли бы проверить положение о том, что Сэди намеревалась обмануть человека, который не делился с ней пищей? Деннетт (Dennett, 1983) предлагает следующее.
Мы создаем для всех шимпанзе совершенно иную ситуацию: они имеют дело с прозрачными пластмассовыми ящиками. Обезьяны должны были понять, что поскольку они - и любой другой - могут видеть через стенки ящика, то и кто-нибудь другой тоже может видеть и, следовательно, узнать, что именно находится внутри ящика. Затем мы можем провести одноразовый эксперимент на новое поведение, в котором будем использовать прозрачный пластмассовый ящик и непрозрачный ящик, а пищу положим в первый из них. Затем входит экспериментатор-конкурент и дает возможность Сэди увидеть, что он смотрит в правильном направлении - на пластмассовый ящик. Если Сэди и в этой ситуации показывает на непрозрачный ящик, то, к сожалению, она действительно не улавливает изощренной идеи обмана. Конечно, и этот эксперимент задуман еще недостаточно совершенно. Прежде всего Сэди могла указать на непрозрачный ящик от отчаяния, не видя лучшего выхода. Чтобы усовершенствовать этот эксперимент, нужно было предоставить обезьяне возможность выбрать еще какой-то вариант поведения, который казался бы ей более подходящим только в том случае, если бы первый вариант поведения она считала безнадежным, как в описанном выше случае. Кроме того, не могла ли Сэди прийти в замешательство от странного поведения экспериментатора-конкурента? А не удивляло ли ее то обстоятельство, что конкурент, не обнаружив пищи там, куда она указала, сразу сел в углу комнаты и начал «дуться», вместо того, чтобы проверить другой ящик? И не должна ли была она удивиться, обнаружив, что ее трюк «сработал»? Ее должно было заинтересовать: неужели ее конкурент-экспериментатор такой бестолковый? Итак, необходимы дальнейшие, гораздо лучше продуманные эксперименты с Сэди и другими существами.
ДЛЯ ЗАПОМИНАНИЯ
1. Невербальная коммуникация наблюдается у человека как часть его повседневного поведения и имеет много общего с коммуникацией животных.
2. Язык нельзя определить как нечто присущее только человеку, поскольку коммуникация животных характеризуется многими чертами, напоминающими язык.
3. Человекообразные обезьяны не могут научиться говорить, но они способны научиться общению с человеком при помощи знаков, символизирующих слова.
4. Согласно представлениям Хомски (Chomsky, 1972), способность человека к языку связана со специфическим типом психической организации, а не просто с более высокой степенью интеллекта.
5. Вопрос о том, является ли когнитивность неотъемлемой частью языка высших обезьян, зависит от того, различают ли они такие категории, как «знание как» и «знание что».
6. Хотя и считается, что в поведении человека важную роль играют психические образы, они, по-видимому, не обязательно необходимы для управления сходными формами поведения у животных.
Рекомендуемая литература
Gardner В. Т., Gardner R. А. (1969). "Teaching sign language to a
chimpanzee", Science, 165, 664-672. Griffin D.R., 1982. Animal Mind - Human Mind, Berlin, Springer - Verlag.
Passing ham R. E., 1982. The Human Primate, New York, Freeman. Rumbaugh D.M., 1977. Language Learning by a Chimpanzee, New
York, Academic Press. Terrace H.S., 1979. Nim, London, Eyre Methuen.
Интеллект, использование орудий и культура
Дарвин считал, что в развитии психических способностей животных существует эволюционная непрерывность; он выступал против широко распространенного мнения о том, что животные представляют собой просто автоматы и по сравнению с человеком находятся на гораздо более низком уровне психического развития. В своей работе «Происхождение человека» (1871) Дарвин утверждал, что «животные обладают определенной способностью к рассудочной деятельности» и что «разница между психикой человека и психикой высших животных, как бы ни была она велика, - это, конечно, разница в степени, а не в качестве». В публикациях ученика Дарвина Джорджа Романеса (Romanes) интеллект животных был даже преувеличен; его книга «Ум животных» (1882) была первой попыткой научного анализа интеллектуальной деятельности животного. Многие доводы Романеса имели анекдотический характер, и его книга была полна историй, рассказанных респектабельными членами Викторианского общества.
Романее определял интеллект как способность «подгонять» свое поведение к изменяющимся условиям жизни. Его некритическая оценка способностей животных вызвала протест со стороны Моргана (Morgan, 1894) и последующих бихевиористов, которые пытались свести интеллект животных только к специфическим способностям. Они критиковали «анекдотический» подход к этой проблеме и разработали строгие критерии, на основе которых можно было говорить о психических способностях животных. Ллойд Морган вдохновил Эдварда Торндайка (Thorndike) на исследование процесса научения методом проб и ошибок (см. гл. 7), и, таким образом, он невольно оказался основателем бихевиористского направления в психологии животных. В своей книге «Введение в сравнительную психологию» (1894) Морган высказал предположение о том, что способности более высокого порядка развиваются на основе более примитивных; он предложил психологическую шкалу психических способностей.
Хотя эта идея об эволюционной шкале способностей в виде некой лестницы и оказала заметное влияние на развитие психологии животных, однако в настоящее время эта точка зрения уже неприемлема. При изучении строения мозга (например, Hodos, 1982) и способностей животных различных видов (например, Macphail, 1982) стало совершенно очевидным, что животные различных видов в различных экологических условиях проявляют огромное разнообразие типов интеллектуальной деятельности. Это очень затрудняет определение понятия интеллект, но подчеркивает важность его исследования у животных с функциональной точки зрения, так же как и изучения обеспечивающих его механизмов.
В этой главе мы рассмотрим некоторые подходы, с помощью которых психологи пытались решить проблему оценки интеллектуальной деятельности животных. Мы достаточно подробно разберем использование орудий как пример поведения, которое большинство людей считает показателем наличия интеллекта; и наконец, мы обсудим проблему передачи культурных традиций как средство, которое дает возможность животным приобретать новые формы поведения.
27.1. Интеллектуальная деятельность животных в сравнительном аспекте
Существуют два основных пути оценки интеллекта животных. Один из них состоит в том, чтобы оценивать поведение, а другой - в том, чтобы изучать мозг. В прошлом оба этих подхода основывались на том, что существует линейная последовательность в развитии от низших, неинтеллектуальных животных, отличающихся сравнительно простым мозгом, к высшим, интеллектуальным животным, мозг которых имеет сложное строение. Обозревая все животное царство в целом, мы, казалось бы, находим подтверждение такого впечатления (см. гл. 11), но когда мы ближе знакомимся с теми или иными особыми случаями, то обнаруживаем здесь много явных отклонений. И это не исключения из общего правила, а следствие того факта, что эволюция шла не линейно, а давала множество разветвлений, на каждом из которых происходит адаптация к своему комплексу внешних условий. Таким образом, животные могут быть весьма сложными в каких-то одних отношениях и достаточно простыми - в других. Вместе с тем животные различных видов могут достигать одинаковой степени сложности, находясь на различных ветвях эволюционного древа.
При сравнении мозга животных различных видов можно ожидать, что между относительным размером какой-то отдельной структуры и степенью сложности поведения, которое регулируется этой структурой, существует определенная связь. Чем больше какое-то животное использует определенную особенность своего поведения в процессе адаптации к окружающей среде, тем больше будет число нейронов и их взаимосвязей в соответствующих областях мозга. Это легко видеть при сравнении специализированных структур мозга, например структур, связанных с различными сенсорными процессами. Гораздо труднее разобраться в том случае, когда приходится рассматривать области мозга более общего функционального назначения, поскольку они могут быть увеличены за счет того, что разные виды животных подвергались различным давлениям отбора (Jerison, 1973).
Многие традиционные идеи в отношении эволюции мозга позвоночных были подвергнуты сомнению. Так, например, вопреки популярным представлениям выяснилось, что в эволюционном ряду рыбы-пресмыкающиеся-птицы-млекопитающие не наблюдается прогрессивного увеличения относительных размеров мозга, а в последовательности миноги-акулы-костистые рыбы-земноводные-пресмыкающиеся-птицы-млекопитающие не наблюдается увеличения относительных размеров переднего мозга (Jerison, 1973). Действительно, относительные размеры переднего мозга у некоторых акул и млекопитающих практически одинаковы (Northcutt, 1981). В течение долгого времени полагали, что конечный мозг акул и костистых рыб прежде всего связан с чувством обоняния, однако теперь считают, что обонятельное представительство в этой области мозга у немлекопитающих животных не больше, чем у млекопитающих (Hodos, 1982). Мысль о том, что низшие позвоночные обладают недифференцированным передним мозгом, также была подвергнута сомнению (Hodos, 1982).
Пытаясь осмыслить наше представление об интеллекте животных в свете современных данных нейроанатомии, Ходос (Hodos, 1982) приходит к следующему заключению: «Если мы сталкиваемся с признаками интеллекта у представителей животного мира и соотносим их со степенью развития нервных структур, мы должны отказаться от линейных, иерархически организованных, моделей, которые преобладают в обоих типах исследования. Нам следует принять более общее определение интеллекта, чем то, которое «привязано» к нуждам и оценкам человека. Мы должны признать тот факт, что история эволюции характеризуется дивергенцией и нелинейностью, и мы не можем ожидать плавных переходов от одного большого таксона к другому. Наконец, мы не можем себе позволить, чтобы наши знания о центральной нервной системе млекопитающих создавали у нас какие-либо предубеждения при поиске нервных коррелятов интеллекта у других классов позвоночных. Если мы не изменим таким образом наше мышление, у нас, по-видимому, останется мало надежды продвинуться хоть немного дальше в наших попытках понять взаимосвязи между психикой человека и психикой животного и соответствующими им нервными субстратами».
Теперь вернемся к вопросу о том, каким образом можно оценить интеллект животного по его поведению. С тех пор как Бине (Binet) в 1905 г. разработал тесты для определения интеллектуального уровня человека, был сделан значительный прогресс в их улучшении и усовершенствовании. Этот прогресс был обусловлен прежде всего тем, что стало возможным дать оценку различным тестам, сопоставляя результаты этих тестов с последующими успехами испытуемых в процессе учебы. Современные тесты по определению коэффициента умственного развития (intelligence quotient - IQ) значительно точнее способны предсказать, насколько далеко продвинется данный человек в области интеллектуальных достижений. Однако остается много трудностей, особенно при попытке сравнивать общий интеллект людей, имеющих различные уровни культуры. Оценивать интеллект животных оказывается гораздо сложнее, поскольку не существует никакого способа проверить обоснованность того или иного теста и поскольку животные различных видов сильно различаются по своим возможностям с точки зрения выполнения той или иной деятельности.
До недавнего времени оценка интеллекта животных в основном базировалась на изучении тех способностей, которые обычно считаются показателем интеллекта у человека. Современный тест для определения IQ включает в себя различные разделы, предназначенные для оценки памяти человека, его арифметических и логических возможностей, способностей к языку и формированию понятий. Как мы уже видели, голуби, по-видимому, обладают удивительной способностью к формированию таких понятий, как вода, дерево и человек. Должны ли мы считать это признаком большого интеллекта? Обсуждая языковые способности животных, мы пришли к заключению, что способности человека в этом отношении намного превосходят аналогичные способности любого животного, даже хорошо обученного.
Но что это означает? Значительное превосходство человеческого интеллекта или его высокую специализированность в плане использования языка?
Для сравнения интеллектуальных способностей животных, относящихся к различным видам, трудно придумать тест, который не был бы предвзятым в том или ином смысле. Многие из прежних тестов для определения способности животного решать какие-то проблемы были ненадежными (Warren, 1973). Порой один и тот же тест, проведенный на животных одного и того же вида, в зависимости от типа используемой аппаратуры давал совершенно различные результаты.
Предпринималось много попыток выяснить, могут ли животные справиться с задачами, которые требуют научения какому-то общему правилу принятия решения. Животных можно научить выбирать из группы предлагаемых предметов такой, который соответствует образцу. Приматы быстро научаются решать такого рода задачу, а голубям для этого требуется гораздо больше попыток. Гарри Харлоу (Harlow, 1949) разработал тест для измерения способности животного следовать каким-либо правилам и делать правильные выводы. Вместо того чтобы проверить обезьян на способность решать какую-то одну задачу простой зрительной дискриминации (рис. 27.1, 4), Харлоу предлагал им последовательно ряд тестов, в которых для решения задачи нужно было следовать одному и тому же правилу. Например, животному можно предложить ряд задач на различение такого типа, как показано на рис. 27 Л,Б. Хотя в каждой задаче использовались другие предметы, правило решения было одинаковым: пищевое вознаграждение в каждом случае (в пределах данной задачи) находится всегда под одним и тем же предметом независимо от того, какое положение он занимает. Если по мере предъявления последовательности таких однотипных задач животное решает их все лучше, то в таком случае говорят, что у него сформировалась установка научения (learning set).
Как видно из рис. 27.1, при исследовании способности животных научиться тому, что общее правило решения является одним и тем же для целого набора задач и что для получения правильного решения нужно руководствоваться одним-единственным принципом, можно использовать различные типы задач. Критики этой методики замечали, что способность животных различных видов формировать установку научения сильно зависит от того, каким образом проводятся тесты (Hodos, 1970). Однако даже с учетом мнения этих критиков следует, по-видимому, признать, что животные различных видов действительно отличаются по своей способности формировать установку научения (Passingham, 1981). Когда разные виды животных ранжировали в соответствии со скоростью улучшения их ответов при последовательном предъявлении однотипных задач, то их ранг можно было угадать на основе индекса развития мозга (Ridell, 1979; Passingham, 1982). С помощью этого индекса оценивается число нервных клеток мозга, избыточных по отношению к тем, которые необходимы для регуляции соматических функций (Jerison, 1973). Итак, создается впечатление, что для оценки интеллекта животных можно разработать тесты, подобные тестам для определения интеллекта человека, и эти тесты позволяют различать психические способности животных различных видов.
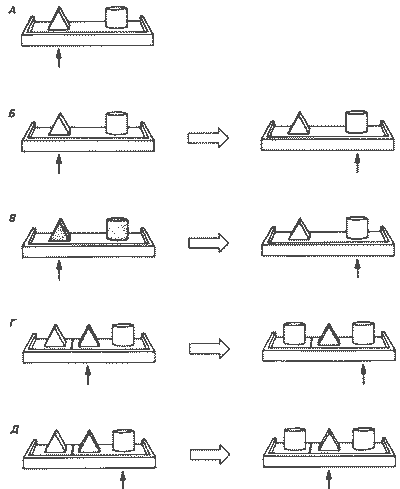
Рис. 27.1. Серии задач на различение, которые были использованы для исследования установки научения. А. Простая дискриминация (стрелкой показан правильный выбор: предмет, под которым находится пища). Б. Обратная задача (решение животного должно быть противоположным тому, которое было правильным в предыдущей задаче). В. Условная задача (нужно выбрать один предмет, если оба предмета серые, и другой, если оба белые). Г. Задача на соответствие (животное должно выбрать предмет, который соответствует образцу, располагающемуся в левой части подноса). Д. Задача на несходство (нужно выбрать тот предмет, который отличается от двух других). (По Passingham, 1981.)
Мнение, что такие тесты представляют собой истинную меру интеллекта, подкрепляется данными о том, что качество выполнения этих тестов коррелирует с показателем размеров мозга. Сходные результаты были получены и при использовании тестов другого типа, представленных на рис. 27.1. Так, например, было показано, что макаки-резусы и шимпанзе в отличие от кошек гораздо быстрее улучшают свои показатели при решении серии задач по различению объектов, если у них был предварительный опыт по решению реверсивных задач, т. е. задач, в которых периодически производилась смена подкрепляемого выбора предмета (Warren, 1974). Эти два типа задач решаются на основе общих принципов, которые способны использовать макаки и шимпанзе, тогда как кошки лишены такой способности. Аналогичные различия между кошками и обезьянами можно отметить и в случае экспериментов с решением задач на несходство, в которых животное должно из группы предметов выбрать непарный (Warren, 1965). Критики этих экспериментов утверждают, что они с неизбежностью проводятся таким образом, что животным одного вида их выполнять легко, а животным другого вида - трудно (Macphail, 1982). Но даже если описанные различия принимать всерьез, они отражают только один аспект интеллектуальной деятельности, и неудивительно, что макаки и высшие обезьяны хорошо выполняют тесты, предназначенные для определения IQ человека, поскольку все они относятся к приматам.
Рис. 27.2. Формирование установки научения зрительной дискриминации у млекопитающих. Процент правильных ответов во второй пробе при решении каждой задачи как функция числа предложенных задач. (По Passingham, 1981.)
27.2. Использование животными орудий
В течение долгого времени способность использовать орудия рассматривали как одно из проявлений интеллекта, а способность изготовлять орудия считали признаком, который отличает человека от всех других животных. Теперь, когда об использовании животными орудий мы знаем гораздо больше, этот вопрос не представляется нам столь ясным, хотя производство орудий по-прежнему рассматривается в основном как важный фактор, который оказал большое влияние на эволюцию человека.
Использование орудий можно определить как использование тех или иных предметов внешнего мира в качестве функционального продолжения какой-либо части тела для достижения ближайшей цели (Lawick-Goodall, 1970). Не все случаи манипуляции животных с различными предметами отвечают этому определению. Например, мы уже знаем, что вороны поднимают в воздух моллюсков и бросают их вниз на камни, чтобы разбить у них раковину. Подобное поведение наблюдается и у других птиц. Так, например, певчий дрозд (Turdus philomelos) берет в клюв улитку и начинает колотить ею по камню, как по наковальне, а вороны (Corvus corax) и некоторые грифы-бородачи (Gypaetus barbatus) сбрасывают вниз кости, чтобы расколоть их и съесть костный мозг. Однако в данном случае камень, используемый животным в качестве наковальни, не считается орудием, поскольку наковальня не является продолжением тела животного.
Известно, что один из грифов, стервятник обыкновенный (Neophron per спор terus), раскалывает яйца африканского страуса, бросая их на камни. Это нельзя считать использованием орудий. Но грифы могут также поднимать камень в воздух и бросать его на гнездо страуса или брать камень в клюв и бросать его на яйцо (рис. 27.3). Такое использование камня уже считается применением орудия, поскольку камень можно рассматривать как продолжение тела грифа.

Рис. 27.3. Гриф готовится бросить камень, чтобы разбить яйцо страуса. [Фотография J. M. Pearson (Biofotos).]
Если животное чешется или трется о дерево, то нельзя считать, что оно использует его в качестве орудия, но если слон или лошадь подберут палку, чтобы почесаться с ее помощью, то будет считаться, что они используют палку как продолжение своего тела для достижения немедленной цели. Однако птица, которая носит веточки для постройки гнезда, использует их как материал, а не как продолжение своего тела. Обычно гнездо не рассматривают как орудие для выращивания птенцов, поскольку оно предназначено для достижения долговременной, а не ближайшей цели.
Галапагосский дятловый вьюрок (Cactospiza pallidd) отыскивает насекомых в трещинах древесной коры, используя для этого колючку кактуса, которую он держит в клюве (рис. 27.4). Согласно приведенному выше определению, это типичный пример использования птицей орудия, однако можно ли считать это признаком интеллектуальной деятельности? С функциональной точки зрения использование колючки кактуса для отыскания пищи - это разумное решение определенной проблемы. О человеке, который находит такое решение этой проблемы, говорят, что он проявляет признаки интеллекта. И те исследователи, которые намерены судить об интеллекте только на основе соответствующих реакций организма на определенные условия, должны допустить, что этот вьюрок ведет себя разумно.
Если бы оказалось, что поведение дятлового вьюрка носит в значительной степени врожденный характер, то мы могли бы уже не считать его пищедобывательное поведение проявлением интеллекта. Наблюдения за молодым дятловым вьюрком, взятым из гнезда в стадии оперившегося птенца, показали, что птица уже в раннем возрасте начинает манипулировать прутиками (Eibl-Eibesfeldt, 1967). Если этому голодному птенцу показать в какой-либо ямке насекомое, то он бросит прутик и будет пытаться достать насекомое клювом. Постепенно птенец начинает доставать насекомых прутиком, и кажется вполне вероятным, что в развитии такого поведения определенную роль играет научение. Но дает ли все это нам больше оснований рассматривать такое поведение как показатель интеллекта? Здесь нам нужно быть весьма осторожными. Даже если в развитии описанной выше активности какую-то роль действительно играет научение, то все же кажется вполне возможным, что дятловые вьюрки оказываются генетически предрасположенными к научению этому конкретному способу манипулировать прутиками почти так же, как некоторые птицы предрасположены к тому, чтобы научиться определенной песне. Напротив, пищедобывательное поведение представляет собой функциональный эквивалент интеллектуального поведения, и мы должны устоять перед искушением, чтобы не сказать, что птица на самом деле не обладает интеллектом только лишь потому, что она - птица, а не млекопитающее. Мы должны с большой осторожностью использовать для оценки поведения птиц те же критерии, которыми мы пользуемся при оценке аналогичного поведения шимпанзе.

Рис. 27.4. Галапагосский вьюрок пользуется колючкой для поиска добычи. (Фотография Alan Root, Survival Anglia, Oxford Scientific Films Limited.)
При наблюдениях за живущими на свободе дикими шимпанзе было обнаружено, что для добывания пищи они пользуются палками, прутиками и стеблями травы. Последние могут использоваться для добывания термитов (рис. 27.5). Обезьяны тщательно выбирают эти стебельки и иногда даже видоизменяют их, чтобы они стали более подходящими для выполнения соответствующей задачи. Например, если кончик стебля загнется, то обезьяна может откусить его (Lawick-Goodall, 1970). Молодые шимпанзе нередко манипулируют стебельками травы во время игры, однако использовать их для добывания пищи они начинают только в возрасте около трех лет. Но даже и тогда они сначала делают это достаточно неуклюже и могут выбирать неподходящие для этой цели орудия. Создается впечатление, что искусству добывания термитов не так легко научиться.

Рис. 27.5. Шимпанзе пользуется веткой, чтобы достать термитов.
Было также обнаружено, что дикие шимпанзе используют палки для добывания меда из пчелиных гнезд и для выкапывания растений со съедобными корнями. Они используют также листья в качестве губки, чтобы добыть питьевую воду из дупла или других более мелких полостей дерева, а также для чистки отдельных частей тела. Вопрос об использовании орудий наиболее интенсивно изучался на диких шимпанзе, однако определенный материал был получен и на других диких приматах. Так, например, павианы используют камни для раздавливания скорпионов и прутики - для добывания насекомых (Kortland, Kooij, 1963).
По всей вероятности, в естественной среде обитания способность использования орудий формируется у каждого отдельного животного в результате как подражательного, так и инструментального научения. В этом отношении использование орудий у приматов трудно отличить от развития пищедобывательного поведения у дятлового вьюрка. Некоторые биологи хотя и допускают, что использование орудии само по себе еще не является признаком интеллекта, однако утверждают, что оно создает предпосылки для истинно разумного поведения, в том числе и для возникновения совершенно новых его форм.
Предположение о том, что новые формы поведения в добывании термитов действительно встречаются у шимпанзе, подтверждается результатами сравнения способов добывания, используемых животными, относящимися к различным популяциям. Так, например, в Гомбе (Восточная Африка) шимпанзе используют прутики, не очищая их предварительно от коры. Иногда они по очереди используют каждый конец прутика. Шимпанзе из Окоробико (Центральная Африка) обычно очищают прутик от коры, перед тем как добывать насекомых, и используют только один конец прутика. Шимпанзе с горы Ассирик в Сенегале (Западная Африка) вообще не пользуются прутиком, а с помощью больших палок проделывают в термитниках отверстия, через которые выбирают термитов рукой (McGrew et al., 1979). Обнаруженные различия позволяют предположить, что в пределах популяции существует определенное число вариаций поведения, которые могут привести к новым формам активности, соответствующим местным условиям. Способу «выуживания» термитов обезьяны научаются путем подражания, и он распространяется в популяции посредством «культурной преемственности».
Предположение о том, - что некоторые формы поведения, типичные для определенной популяции, поддерживаются в ней за счет культурной преемственности, вполне подтверждается данными, приводимыми в научных публикациях (Bonner, 1980; Mundinger, 1980). В нескольких (хотя и немногих) случаях удалось наблюдать возникновение у животных новых форм поведения и зарегистрировать, каким образом это поведение распространялось среди членов популяции. Хорошо известен случай с японскими макаками (Macacafuscatd), обитающими на острове Кошйма (Kawamura, 1963). Чтобы заставить обезьян держаться на открытом месте, где за ними легче было бы наблюдать, экспериментаторы дополняли их меню, разбрасывая по берегу «клубни» батата. Они видели, как 16-месячная самка по кличке Имо отмывала в ручье песок с «клубней». Она регулярно проделывала эту операцию, и вскоре ей стали подражать другие обезьяны, особенно ее сверстницы. В течение 10 лет эта привычка распространилась почти во всей популяцииза исключением только взрослых особей старше 12 лет и молодняка, не достигшего еще годовалого возраста. Два года спустя Имо придумала еще одну операцию по очистке пищи. Экспериментаторы разбросали по берегу зерна злаков, и обезьяны собирали их по одному. Имо же набрала полную горсть смешанных с песком зерен и бросила все это в воду. Песок пошел ко дну, а зерна оказалось легко собрать с поверхности воды. Эта новая операция по очистке пищи распространилась среди особей популяции точно так же, как и способ мытья «клубней» батата. Новое поведение прежде всего переняли сверстницы Имо. Матери научились этой операции от молодых обезьян, а взрослые самцы освоили ее последними.
Была ли Имо особенно умной обезьяной? С одной стороны, можно утверждать, что ее научение было схоже с так называемым научением типа инсайта, описанным Кёлером, о котором говорилось в гл. 19. Имо сделала свои открытия случайно и научилась их использовать. Совсем необязательно, чтобы в этой ситуации у нее имел место какой-то особый инсайт. С другой стороны, многие изобретения человека возникают подобным же образом. Если какая-либо изобретательность в поведении приводит к истинному улучшению условий жизни обезьян, то это расценивается как форма адаптивного поведения, приобретенная посредством усилий индивидуальной особи. Если судить по результатам, то Имо представляется высокоинтеллектуальной обезьяной. Но коль скоро мы считаем, что интеллект включает в себя такой механизм, как рассуждения, то нам необходимо больше знать о мыслительных процессах Имо, прежде чем мы сможем сделать какой-либо вывод.
27.3. Культурные аспекты поведения
Эволюция представляет собой результат естественного отбора, и наследование приобретенных свойств обычно невозможно. Как бы хорошо ни было адаптировано отдельное животное к условиям своей среды обитания - путем научения или в результате физиологических адаптации, - эти приобретенные адаптации не могут быть переданы потомству с помощью генов. Именно эта точка зрения сейчас широко принята среди биологов. Однако, как мы уже знаем, информация может передаваться от родителей к потомству путем подражания и посредством импринтирования. Передачу информации от одного поколения к другому негенетическим путем называют культурной преемственностью.
Мы уже знаем (гл. 20), что благоприятные для научения чувствительные периоды у многих животных проявляются на раннем этапе их жизни. Именно во время этих периодов молодняк часто перенимает навыки от своих родителей. Так, например, белоголовая воробьиная овсянка запомнит песню своих родителей только в том случае, если услышит ее во время чувствительного периода своего развития между 10 и 50 сут жизни. Если во время этого периода отдельные птенцы были лишены возможности услышать песню своего вида, то в течение всей своей дальнейшей жизни они уже никогда не могли научиться специфической песне белоголовой воробьиной овсянки. Следует сказать, что птенцы этого вида овсянок никогда не научаются песням, которые бы существенно отличались от их собственной, тогда как другие птицы, например снегирь, способны научиться петь песни совершенно других видов. Так, воспитанный канарейкой снегирь заимствует ее песню. Тенденция копировать песню родителей приводит к возникновению региональных вариаций даже у тех видов, которые способны освоить только песни, напоминающие песни их вида. К примеру, у обитающих в районе Сан-Франциско популяций белоголовой воробьиной овсянки, которые были разделены всего лишь несколькими километрами, обнаружены различные диалекты (Marler, Tamura, 1964).
Диалекты животных представляют собой элементарную форму явления, называемого традицией. Птенцы белоголовых воробьиных овсянок обязательно слышат пёсенные диалекты, характерные для тех мест, где они родились, поскольку чувствительные периоды научения пению приходятся у них на тот период развития, когда они еще не способны покинуть гнездо. Еще одна форма традиционного поведения - это миграции по определенным маршрутам некоторых млекопитающих и птиц. Гуси, утки и лебеди совершают эти миграционные перелеты стаями, состоящими из взрослых и молодых птиц. Молодняк учится у взрослых лететь по маршруту, характерному для данной популяции, останавливаться в традиционных местах отдыха, использовать места размножения и зимовки (Schmidt-Koenig, 1979). Существует и много других примеров. Северные олени также проявляют верность традиционным маршрутам миграции и тем районам, где у них обычно рождаются оленята. Мигрирующие лососи появляются на свет в пресноводных ручьях, а затем уплывают в море. Молодые рыбы импринтируют запах воды родного ручья и возвращаются к нему, когда становятся взрослыми, чтобы нереститься в традиционном месте. Известно, что некоторые тропы олени и другие млекопитающие используют в течение столетий. В некоторых районах Великобритании и Германии, где дороги пересекают традиционные пути миграции жаб, служители заповедников во время брачного периода этих земноводных стоят на страже, чтобы предотвратить их гибель под колесами машин.
Простые формы традиционного поведения не требуют от животных каких-либо особых способностей к научению или особого их обучения. Это поведение проявляется как неизбежное следствие условий, в которых воспитываются молодые животные, и их стремление импринтировать все особенности местообитания, родителей и своих сверстников. Однако у некоторых животных поведение формируется на основе более сложного научения. Мы уже видели, что некоторые формы использования орудий у приматов могут распространяться по всей популяции - по-видимому, в результате подражательного научения.
Однако подражание не является необходимым признаком высокого интеллекта. Животные могут копировать друг друга в результате простого социального облегчения (facilitation) проявлений соответствующей активности. Многие животные, питаясь в группе, съедают больше, чем при питании в одиночку. Это было продемонстрировано в экспериментах на цыплятах, щенках, рыбах и опоссумах. Если цыпленку дают возможность наесться досыта, а затем подсаживают его к другим цыплятам, которые продолжают есть, то сытая птица тоже начнет есть. Замечено также, что домашние цыплята стремятся клевать, когда это делают другие цыплята, даже если поблизости нет никакой пищи. Цыплята стараются клевать любые мелкие объекты такого типа, которые клюет их мать. На воробьях и зябликах было продемонстрировано стремление животных поедать именно ту пищу, которую потребляют другие (Davis, 1973).
Социальное облегчение определенного поведения можно обнаружить и при избегании животными вредной пищи. Предпринятые в США попытки уничтожить большие стаи обыкновенных ворон с помощью ядовитых приманок не привели к успеху, поскольку большая часть птиц начинала избегать этих приманок после отравления нескольких птиц. Подобно этому реакция всего лишь одной крысы на новую пищу может быть достаточной, чтобы определить реакции других крыс группы. Если одна крыса ест данную пищу, то другие присоединяются к ней, но если она, обнюхав пищу, отвергает ее, то другие тоже от нее отказываются. Иногда первая крыса мочится на эту приманку и таким образом предупреждает других о том, что ее следует избегать (Rozin, 1976b).
Очень часто привлечение к пище или, напротив, отвергание ее за счет социальных взаимодействий возникает как результат стремления животных изучить места, где побывали другие особи вида, а не как результат прямого копирования поведения других. По-видимому, именно таким образом объясняется случай традиционного воровства молока, которое совершали в Англии синицы-лазоревки. В течение многих лет в Англии к домам жителей ранним утром доставляли молоко и оставляли его у порога. Лазоревки, а иногда и большие синицы, проклевывали сделанные из фольги бутылочньге крышки и угощались собравшимися в верхней части бутылок жирными сливками (рис. 27.6). Хайнд и Фишер (Hinde, Fisher, 1951) сообщают о том, что сначала случаи такого воровства появились в отдельных районах страны, а затем постепенно оно распространилось и дальше, что наводит на мысль о том, что птицы научаются такого рода навыкам друг у друга.

Рис. 27.6. Синица-лазоревка открывает бутылку, чтобы полакомиться молоком. (Фотография John Markham. С разрешения Bruce Coleman Ltd.)
Другой интересный пример традиционного пищевого поведения наблюдал Нортон-Гриффитс у молодых куликов-сорок (Norton-Griffiths, 1967; 1969) - птиц, которые живут на морском берегу и питаются мидиями, т.е. моллюсками, заключенными в твердую двустворчатую раковину. Взрослые птицы пользуются одним из двух способов добычи мяса этого моллюска. Либо они очень сильно ударяют клювом, как молотком, по самому тонкому участку раковины, либо вставляют клюв в открытый сифон, когда мидии находятся под водой, и разрезают им приводящую мышцу, которая держит обе створки вместе, закрывая раковину. Взрослые птицы вскрывают раковины одним или другим способом, и никогда одна и та же птица не пользуется обоими способами вскрытия раковины. Более того, оба члена брачной пары используют один и тот же метод добычи мяса мидии. Меняя местами яйца тех и других птиц, Нортон-Гриффитс показал, что птенцы научаются способу вскрытия раковин мидий от своих родителей. В раннем возрасте у всех птенцов наблюдаются движения, которые соответствуют приемам и тех и других родителей, но птенцы, по-видимому, не практикуют их. Вполне вероятно, что оба способа добычи мяса до определенной степени врожденные. Каким образом молодые птицы приобретают необходимый им навык, остается до конца не выясненным. Они живут со своими родителями 18-26 нед, тогда как птенцы тех куликов-сорок, которые ловят главным образом другую добычу, остаются с родителями всего лишь 6-7 нед. Птенцам, воспитанным в традициях питания мидиями, приходится принимать пищу от родителей, пока они находятся в поре ученичества. Быть может, птенцы научаются добывать себе пищу путем наблюдения, а быть может, дело в том, что для такого способа добывания необходима координация движений, требующая для своего формирования длительного времени.
Из этих примеров становится очевидно, что преемственность культурных традиций не обязательно требует большого интеллекта от отдельных особей данной популяции. Хотя и высокоразвитая культура, и высокоразвитый интеллект - это очень важные человеческие особенности, они не обязательно связаны причинной связью. У примитивных народов существует много культурных традиций, которые носят адаптивный характер, однако на первый взгляд не кажутся рациональными. Например, большая часть традиционных методов приготовления блюд из кукурузных зерен туземцами Нового Света основана на том или ином варианте их щелочной обработки. Часто кукурузу варят в течение примерно 40 мин в воде, содержащей древесную золу, щелок или растворенную известь, а затем все это сразу съедают или превращают в тесто, лепешки и т.д. Эта щелочная обработка проводится по причинам чисто традиционным, хотя некоторые и утверждают, что после такой обработки пища становится более вкусной. Однако обработка пищевых продуктов щелочью имеет важное значение для питания. В местной кукурузе содержится мало лизина - аминокислоты, относящейся к группе незаменимых аминокислот. Большая часть содержащегося в кукурузе лизина находится в составе неперевариваемого белка, называемого глютелином. В результате щелочной обработки происходит расщепление глютелина, и при этом выделяется лизин, который значительно улучшает питательные качества пищи. Оказалось, что все туземцы, у которых основу питания составляет кукуруза, используют те или иные способы щелочной обработки пищи. Вполне вероятно, что естественный отбор, действуя через неправильное питание, элиминирует ту часть популяции, которая не следует традициям в обработке пищи.
Таким образом, мы видим, что, хотя культурная преемственность и может действовать в обход биологической наследственности и приводить к очень быстрой эволюции, продукты культурной эволюции по-прежнему остаются объектом естественного отбора. Хотя некоторые склонности к научению и являются существенными для изменения поведенческой культуры, большого интеллекта для этого не требуется, и те люди, которые следуют традиционным видам деятельности, часто оказываются не в состоянии дать рационального объяснения своему поведению.
ДЛЯ ЗАПОМИНАНИЯ
1. Очень трудно сравнивать интеллектуальные способности животных различных видов, поскольку многие из них обладают специфическими навыками и в то же время специфическими «неспособностями» решать какие-то определенные задачи. Следует ли судить об интеллектуальных возможностях животных по типу задач, которые они могут решить, или по тому способу, которым они это делают?
2. Способность к использованию орудий широко распространена в животном мире, а некоторые виды могут даже сами создавать свои собственные орудия. Следует ли рассматривать эту способность как показатель разумной деятельности?
3. Преемственность культурных традиций, посредством которой происходит передача информации от одного поколения к следующему негенетическим путем, наблюдается у многих видов животных. Усвоение традиций может осуществляться в результате импринтирования или подражательной деятельности.
Рекомендуемая литература
Banner J. Т., 1980. The Evolution of Culture in Animals, Princeton,
New Jersey, Princeton University Press. Jerison H. J., 1973. Evolution of the Brain and Intelligence, New York, Academic Press. Macphail E.M., 1982. Brain and Intelligence in Vertebrates, Oxford, Clarendon Press.
Сознание и эмоции у животных
Эмоции - это существенная часть человеческих переживаний. С точки зрения здравого смысла можно прийти к единому мнению о том, что именно подразумевается под эмоциями, и мы обычно единодушны в понимании атрибутов различных эмоций. Однако при изучении эмоций с научной точки зрения возникает целый ряд трудных проблем.
Эмоции могут проявляться на субъективном, физиологическом и поведенческом уровнях, которые трудно согласуются друг с другом. С точки зрения субъективных проявлений эмоции - это, в сущности, субъективные переживания. Не существует такого способа, который помог бы нам узнать, на что именно похожи эмоциональные переживания другого человека. Мы склонны предполагать, что эти переживания точно такие же, как и наши собственные, но у нас нет никакого логического пути, чтобы убедиться в этом предположении. Когда же мы подходим к проблеме эмоционального переживания животных, то здесь мы сталкиваемся с еще большими трудностями. Мы склонны предположить, что похожие на нас животные, например приматы, способны на такие же эмоциональные переживания, как и мы, а животные, которые на нас непохожи, например насекомые, если и способны на какие-либо переживания, то, по всей вероятности, только на те, которые очень отличаются от наших. Однако это всего лишь голос здравого смысла, а не научный вывод. С точки зрения науки, мы не можем утверждать, что у животных имеются какие-то особые субъективные чувства - логически мы вправе делать такие предположения только в отношении других людей.
С точки зрения физиологии эмоциональные состояния человека обычно сопровождаются изменениями вегетативных показателей, но эти показатели не дают нам надежного ключа для определения конкретных эмоциональных состояний. У животных особенно трудно дифференцировать эмоции (например, страх, агрессивность, сексуальные ощущения), даже когда такие физиологические показатели, как повышение частоты сердечных сокращений или нарушение гормонального равновесия, действительно свидетельствуют об эмоциональном возбуждении организма. Другими словами, большинство животных проявляют одни и те же физиологические реакции безотносительно к тому, отражает ли их эмоциональный ответ реакцию страха, агрессии или проявление полового чувства. Таким образом, хотя мы и можем кое-что выяснить на основе физиологических исследований, интерпретация физиологических ответов оказывается весьма трудной.
Чарлз Дарвин (1872) подчеркивал коммуникативный аспект эмоций. Как мы уже видели в гл. 22, он утверждал, что мимические реакции и другие поведенческие признаки эмоций произошли от защитных реакций и других утилитарных аспектов поведения. Хотя эволюционная концепция Дарвина, которую развивали и широко распространяли ранние этологи, в настоящее время считается общепринятой, его представления об эмоциях кажутся весьма примитивными. Дарвин и его ученик Джордж Романее не смущались, когда обозначали эмоции животных терминами, характеризующими эмоции человека. Они писали, в частности, что поведение и мимика провинившейся собаки выражают «стыд» (Darwin, 1872), рыба переживает «ревность», а у попугая проявляется чувство «гордости», когда он произносит свои «слова» (Romanes, 1882). Столь антропоморфический подход вызвал у психологов протест. Например, Морган (Morgan, 1894) пропагандировал такой подход к изучению данной прблемы, в котором не было бы каких-либо спекуляций по поводу мыслей и чувств животных.
В этой главе мы попытаемся понять внутреннюю жизнь животных с точки зрения их субъективного мира, а также их физиологических и поведенческих проявлений. Кроме того, мы обсудим вопрос о том, какое значение имеют эти исследования для обеспечения благосостояния животных и лля выбора условий их содержания.
28.1. Самосознание у животных
На протяжении почти 75 лет нашего столетия прочно господствовало представление бихевиористов о том, что субъективные психические переживания животных не могут быть предметом научных исследований. В течение этого времени такие ученые, как Толмен (Tolman, 1932), оспаривали эту точку зрения, но они практически никак не влияли на господствующее мировоззрение (см. обзор Griffin, 1976). Позиция бихевиористов с логической точки зрения кажется неприступной, однако ее можно обойти различными путями. Один из аргументов заключается в том, что, хотя мы и не можем доказать, что животные обладают субъективными переживаниями, вполне возможно, что на самом деле так оно и есть. А если бы это было так, то что тогда изменилось бы? Другой подход базируется на утверждении, что с эволюционной точки зрения представляется маловероятным, чтобы между человеком и животными в этом отношении был существенный разрыв.
Гриффин (Griffin, 1976), который одним из первых начал планомерную атаку на позиции бихевиоризма, использовал оба этих аргумента. По его мнению, изучение коммуникации животных с наибольшей вероятностью должно принести нам доказательства того, что «они обладают психическими переживаниями и сознательно общаются друг с другом». Однако при исследовании языковых способностей животных в последние годы это давнее обещание оказалось невыполненным. До сих пор вызывает спор поведение шимпанзе, которых обучили некоторым особенностям языка человека, и существует сомнение в том, что когда-либо эти эксперименты позволят нам многое узнать о субъективных переживаниях этих животных (Terrace, 1979; Ristau, Robbins, 1982). Были предприняты самые различные попытки исследовать субъективный мир животных другими способами, к описанию которых мы и переходим.
Осознают ли животные себя в том смысле, что имеют ли они представления о позах, которые принимают, и о действиях, которые совершают? Конечно, поступающая от суставов и мышц сенсорная информация направляется к мозгу, и поэтому животное, по-видимому, должно быть осведомлено о своем поведении. В экспериментах, направленных на выяснение этого вопроса, крыс обучали нажимать на один из четырех рычагов в зависимости от того, какой из четырех активностей было занято животное, когда раздавался звук зуммера (Beninger et al., 1974). Например, если этот сигнал заставал крысу в тот момент, когда она чистила шерсть (was grooming), она должна была нажимать на «груминговый» рычаг, чтобы получить пищевое подкрепление. Крысы научились нажимать на различные рычаги в зависимости от того, занимались ли они чисткой шерсти, ходили, поднимались на задние лапки или находились в покое в\юмент, когда слышался звук зуммера. Результаты подобных экспериментов (Morgan, Nicholas, 1979) показали, что крысы способны строить свое инструментальное поведение на основе информации об их собственном поведении и сигналов, поступающих из внешней среды. В каком-то смысле крысы должны знать о своих действиях, но это совсем не означает, что они их осознают. Они могут осознавать свои действия точно так же, как и внешние сигналы.
Многие животные реагируют на зеркало так, как-будто они видят других особей своего вида. Однако некоторые данные свидетельствуют о том, что шимпанзе и орангутаны могут узнавать себя в зеркале.

Рис. 28.1. Шимпанзе Вики имитирует свое изображение на фотоснимке. (Рисунок по фотографии.)
Молодые шимпанзе, рожденные на воле, пользовались зеркалом, чтобы чистить те части своего тела, которые другим путем увидеть невозможно. Гэллап (Galiup, 1977; 1979) наносил небольшие пятнышки красной краски на бровь и противоположное ухо нескольким шимпанзе, когда они находились под легкой анестезией. По утверждению экспериментатора, шимпанзе, выйдя из наркотического состояния, прикасались к этим частям своего тела не чаще, чем обычно. Тогда он дал обезьянам зеркало. Шимпанзе начали разглядывать свои отражения в зеркале и постоянно трогать окрашенные брови и уши.
Можно ли считать, что способность животного реагировать на какие-то части своего тела, которые оно видит в зеркале, свидетельствует о его самоосознании? Этот вопрос непосредственно связан с более широким вопросом. Свидетельствует ли способность животного подражать действиям других о его «знании себя»? Шимпанзе невероятно искусны в подражании друг другу и людям. Хотя истинное подражание следует очень тщательно отличать от других форм социального научения (Davis, 1973), мало кто сомневается в том, что приматы способны к подражанию. Например, шимпанзе Вики, воспитывавшейся в семье Хейесов (Hayes), было предложено скопировать серию из 70 движений. Многие из этих движений она никогда ранее не видела, но десять из них скопировала сразу, как только ей их показали. Вики научилась производить в ответ на соответствующие демонстрации 55 двигательных актов (рис. 28.1). Она также научилась выполнять довольно сложные домашние дела, например мыть посуду или вытирать пыль (Hayes, Hayes, 1952). Многим из этих действий она подражала спонтанно, без чьих-либо наводящих посылок. Однако шимпанзе по подражательным способностям не смогла справиться с ребенком. Исследователи считали, что подражательная активность Вики соответствует способностям детей в возрасте от 12 до 21 мес. Способность к подражанию иногда считают признаком интеллекта, однако этот тезис стоит взять под сомнение, поскольку подражание наблюдается и у очень маленьких детей, и у самых различных немлекопитающих животных. При изучении пения птиц оказалось, что у многих видов птиц при научении пению наблюдаются некоторые формы подражания звукам, причем некоторые птицы в этом отношении особенно искусны. Попугаи и индийские скворцы майны способны необычайно точно воспроизводить звуки человеческого голоса (Nottebohm, 1976).
Чтобы иметь возможность подражать, животное должно получить внешний слуховой или зрительный пример для подражания и добиться соответствия ему с помощью определенного набора своих собственных моторных инструкций. Например, ребенок, который подражает взрослому, высовывая язык, должен как-то ассоциировать вид языка со своими моторными инструкциями, необходимыми для того, чтобы самому высунуть язык. Ребенок при этом совершенно не обязан знать, что у него есть язык, - он просто должен связать данное сенсорное восприятие с определенным набором моторных команд. Каким образом это происходит, остается загадкой. Однако вопрос о том, необходимо ли самоосознание для осуществления подражательной деятельности, является спорным.
Отчасти проблема состоит в том, что нам необходимо выяснить, что же именно мы подразумеваем под термином самоосознание (selfawareness). Как отмечал Гриффин (Griffin, 1982), многие философы проводят различия между понятиями «осознание», или «знание себя» (awareness), и «сознание» (consciousness. Осознание - это вид восприятия, тогда как сознание включает в себя особый род самоосознания, которое не ограничивается простой осведомленностью о разных частях своего тела или процессах, протекающих в мозгу. Сознание, с этой точки зрения, включает в себя какое-то предположительное знание того, что Я испытываю ощущения или думаю, что это Я - существо, знающее об окружающем мире. Мы разобрали несколько примеров того, что животные обладают знаниями в сфере восприятия, т.е. знают непосредственно воспринимаемые характеристики объектов. Однако способность животного сообщать о своих действиях, подражать действиям других или узнавать свое изображение в зеркале не обязательно требует наличия сознания в том смысле, как оно было здесь определено.
Рассогласование между осознанным и неосознанным восприятиями можно наблюдать у людей с повреждениями мозга. Некоторые люди, у которых повреждены определенные области мозга, связанные с обработкой зрительной информации, сообщают о том, что они частично ослепли. Они не в состоянии назвать объекты, которые предъявляют им в определенных областях поля зрения. Они утверждают, что не могут увидеть эти объекты; однако, когда их просят указать на них, они зачастую могут сделать это очень точно (Weiskrantz, 1980). Один больной точно угадывал, какую линию ему показывали; горизонтальную или диагональную, хотя он и не знал, видит ли он что-либо (Weiskrantz et al., 1974). Это явление, названное слепым зрением, возникает в результате повреждения тех областей мозга, которые ответственны за опознание зрительных сигналов (visual awareness), тогда как другие области мозга, участвующие в зрительном процессе, остаются интактными. Именно эти области мозга помогают больному делать правильное суждение, хотя он и не знает о том, что он видит.
28.2. Физиологические аспекты эмоций
Эмоциональные состояния сопровождаются усилением активности вегетативной нервной системы, отдела нервной системы позвоночных, который регулирует работу внутренних органов, например кровеносных сосудов, сердца, кишечника, легких и некоторых желез. Вегетативная нервная система находится под контролем головного мозга и обеспечивает два типа иннервации, оказывающие противоположные влияния на внутренние органы. Симпатическая нервная система активируется в условиях стресса или напряжения и обеспечивает мобилизацию организма в чрезвычайных обстоятельствах. Ее влияние проявляется в увеличении кровоснабжения мышц, мозга, сердца и легких, усилении частоты сокращений сердца и ослаблении притока крови к кишечнику и периферическим частям тела. Парасимпатическая нервная система анатомически отделена от симпатической; она обеспечивает восстановительную функцию, возвращая к норме кровоснабжение органов и нейтрализуя другие эффекты симпатической активности. Эмоциональное возбуждение сопровождается симпатической активацией.
У человека это проявляется в увеличении частоты сокращений сердца, усилении потоотделения и изменении периферического кровообращения, в результате чего лицо бледнеет или, напротив, становится красным.
Психолог Джеме высказал предположение, что субъективные переживания эмоций берут свое начало в возбуждении сенсорных рецепторов, вовлекающихся в данную эмоциональную реакцию. Например, пугающий раздражитель должен вызвать определенные поведенческие и физиологические изменения, а уже эти изменения порождают субъективное переживание испуга. Как пишет Джеме (James, 1980), когда мы встречаемся с медведем, мы убегаем. Мы убегаем не потому, что пугаемся, - мы пугаемся потому, что бежим. Современник Джемса-Ланге (Lange) предложил схожее объяснение для эмоционального переживания, и эта точка зрения известна в настоящее время как теория Джемса-Ланге.
Кэннон (Cannon, 1927) утверждал, что симпатическое возбуждение во многом характеризуется одинаковой картиной независимо от того, какую эмоцию переживает организм в данный момент. Он показал, что у кошек даже после устранения всех видов сенсорной обратной связи к вегетативным нейронам продолжают проявляться такие признаки эмоционального поведения, как шипение и слюноотделение. Кэннон пришел к заключению, что активация вегетативной нервной системы не является необходимой для эмоционального переживания, хотя эмоциональное переживание и влияет на вегетативную активность. Когда испытуемым вводят гормон адреналин, у них можно наблюдать самые различные симпатические реакции, например потоотделение, усиление частоты сердечных сокращений и т. д. Однако при этом испытуемые не сообщают о каких бы то ни было эмоциональных переживаниях. Некоторые из них говорят об определенных физических симптомах, а другие - о том, что они ощущают «холодную» эмоцию - состояние, напоминающее раздражение или страх (Landis, Hunt, 1932).
Активация вегетативных функций организма может обеспечить основу для эмоциональных переживаний, но она не создает никакой дифференциации в состояниях организма при различных эмоциональных ситуациях. Однако определенная ситуация, будучи воспринятой субъектом, может привести его к интерпретации вегетативного возбуждения в виде соответствующего эмоционального переживания. Эту гипотезу предложили Шехтер и Сингер (Schachter, Singer, 1962). Они вводили испытуемым адреналин,говоря,что это инъекция витамина. Некоторым из подопытных лиц они сказали, что этот укол вызовет у них усиление сердцебиения, покраснение и т. д. (т. е. реальные эффекты адреналина), тогда как другим сообщали, что у них могут возникнуть такие побочные эффекты, как зуд или онемение (т.е. они были дезинформированы). Затем всех испытуемых попросили посидеть в комнате ожидания, перед тем как у них проверят зрение. В этой комнате за ними наблюдал скрытый от них экспериментатор, а другой экспериментатор-ассистент находился вместе с ними. В одном опыте этот второй экспериментатор вел себя таким образом, как будто он был сердит и молчалив, а в другом, напротив, он был весьма несерьезным и веселым. Когда испытуемых попросили оценить их эмоциональные переживания после пребывания в комнате ожидания, оказалось, что между этими тестовыми ситуациями обнаружились значительные различия. Испытуемые, которые были дезинформированы в отношении последствий инъекций, чувствовали себя в присутствии игривого ассистента более веселыми, чем испытуемые, которые были информированы правильно. В присутствии же угрюмого соучастника они чувствовали себя раздраженными и более обиженными, чем контрольные испытуемые. Хотя эти эксперименты и критиковались некоторыми авторами (например, Plutchik. Ax, 1970), тем не менее они дают основание предположить, что в формировании нашего эмоционального переживания принимают участие и ощущение нашего физиологического состояния, и наша оценка внешней обстановки.
Пытаясь оценить субъективные переживания животных с точки зрения физиологии, можно провести прямые сопоставления с человеком, но ни одно из них не будет вполне удовлетворительным. Например, способны ли животные ощущать боль? Когда животных подвергают раздражениям, вызывающим боль у человека, у них наблюдаются физиологические реакции, идентичные реакциям человека. Блокирующие боль вещества, которые ослабляют субъективные, физиологические и поведенческие реакции на боль у человека, оказывают сходные влияния на типичные поведенческие признаки боли у животных. Однако интерпретация таких данных основывается главным образом на использовании поведенческих показателей. Например, просто рефлекторное отдергивание, по-видимому, не может быть хорошим показателем боли (как субъективного переживания), поскольку в мире животных такие реакции весьма распространены и встречаются у очень примитивных организмов. Даже по такому критерию, как крик, трудно оценивать боль. Если собаки и обезьяны в случае тяжелого ранения будут вопить от боли, то антилопа останется относительно молчаливой даже тогда, когда хищник рвет ее на куски. Эти вопли могут служить для того, чтобы получить помощь от других членов группы, но ведь они могут также навлечь опасность на тех, кто на них откликнется.
Чтобы оценить вероятность того, что и животным присущи субъективные переживания, аналогичные переживаниям человека, иногда прибегают к сравнительному анализу анатомии мозга. В качестве показателя прогрессивного развития от простого животного к более интеллектуальному обычно используются весьма простые критерии, такие, как размеры всего головного мозга или корковой его части. Однако результаты тщательных аналитических исследований показали, что в различных группах животных некоторые структуры мозга существенно различаются. Ходос (Hodos, 1982) предположил, что некоторые из этих специализированных областей могут принимать участие в обеспечении определенного типа психической активности, которую у человека обычно связывают с деятельностью коры больших полушарий. Кроме того, при оценке интеллектуальных способностей животных на основе особенностей их мозговых структур часто бывает трудно предложить хороший метод, который не был бы связан с антропоморфическими предположениями (Macphail, 1982).
28.3. Сознание и ощущение страдания
Проблема сознания животных таит в себе много трудностей. Спектр научных представлений по этому поводу очень широк. Одни ученые уверены в том, что сознания у животных нет, а другие утверждают, что у большинства животных сознание есть. Существуют исследователи, считающие, что сознание не может быть предметом научного изучения, и исследователи, считающие эту тему не заслуживающей внимания. Ситуация осложняется еще и тем, что очень трудно прийти к приемлемому определению сознания.
Гриффин (Griffin, 1976) определяет сознание как способность организма создавать психические образы и использовать их для управления своим поведением. Это очень напоминает определение, которое предлагает Оксфордский толковый английский словарь. Быть в сознании - это значит «знать, что ты сейчас делаешь или собираешься делать, имея перед собой цель и намерение своих действий» (см. Griffin, 1982). Согласно Гриффину (Griffin, 1976), «намерение включает в себя психические образы будущих событий, причем намеревающийся представляет себя одним из участников этих событий и производит выбор того образа, который он попытается реализовать». Хотя Гриффин и другие исследователи (например, Thorре, 1974) рассматривали намерение и сознание как неотъемлемую часть одного и того же явления, эта точка зрения не является общепринятой. В гл. 26 было показано, что преднамеренное поведение не всегда требует сознания.
Как мы уже видели раньше, многие исследователи полагают, что сознание нельзя сводить только к знанию своих чувственных восприятии. Например, Хамфри (Humphrey, 1978) понимал сознание как самознание (self-knowledge), которое используется организмом, чтобы предсказать поведение других индивидуумов, а Хаббард (Hubbard, 1975) полагал, что оно подразумевает осознание себя как чего-то отличного от других. Такое знание может быть использовано как основа коммуникации, но это не означает, что сознание непременно включает в себя язык. Мы можем согласиться с Пассингэмом (Passingham, 1982), что «говорящий язык революционизировал мысль. Использование языка для мышления создало условия для того, чтобы интеллект мог достичь гораздо более высокого уровня. Животные думают, но люди способны думать абсолютно по-другому, используя совершенно другой код». Несомненно, что вторжение языка изменило сам способ, каким мы осмысливаем самих себя. Нам трудно представить себе сознание без языка. Однако это не дает нам права считать, что животные, которые не имеют языка или обладают очень примитивным языком, не имеют сознания. Мы уже видели раньше, что у животных, которые не имеют языка, эквивалентного человеческому, можно обнаружить признаки самоосознания. Поэтому мы не должны приравнивать язык к сознанию.
Могут ли животные испытывать осознанное страдание? Если стоять на позиции здравого смысла, то мы склонны предположить, что могут. Когда мы находимся в бессознательном состоянии, мы не страдаем от боли или душевных мук, поскольку какие-то области нашего мозга оказываются инактивированными. Мы не знаем, однако, отвечают ли эти области только за сознание или же за сознание плюс еще какие-то аспекты работы мозга. Таким образом, хотя в бессознательном состоянии мы не испытываем боли, мы не можем на основе этого сделать вывод о том, что сознание и страдание идут рука об руку. Вполне может быть, что все то, что лишает нас сознания, одновременно прекращает ощущение боли, но одно и другое не имеют причинной связи.
У нас нет никакой концепции в отношении того, что может включать в себя совокупность сознательного опыта животного, если таковой существует. Поэтому мы не можем сделать никакого заключения о том, существует ли какая-либо связь между сознанием животных и их чувством страдания. В своем неведении мы, должно быть, очень неправы, когда, думая о животных, полагаем, что ощущение страдания может быть лишь у тех из них, которые обладают интеллектом, языком и у которых обнаруживаются признаки осознанных переживаний.
Есть свои недостатки и свои достоинства в том, что мы используем самих себя в качестве моделей, на которых пытаемся изучить возможности ощущений у животных (Dawkins, 1980). Слишком слаба научная основа для проведения аналогии между психическими переживаниями человека и животных. Было бы некорректно с научной точки зрения приходить к какому-то заключению о психических переживаниях животных на основе таких данных. Вместе с тем мы сами делаем заключения о психических переживаниях других людей только на основе аналогии с нашими собственными переживаниями. Когда мы видим, как другой человек страдает или кричит от боли, мы не пренебрегаем этим, хотя и не можем доказать идентичность его психических переживаний с нашими. Мы «истолковываем сомнения в пользу обвиняемого» и приходим к нему на помощь. Быть может, в отношении представителей других биологических видов мы тоже должны истолковывать наши сомнения в их пользу?
28.4. Перспективы эволюции
Нам кажется естественным помогать другим людям, когда они испытывают страдания. Наши симпатии, по всей вероятности, имеют врожденную основу, хотя в какой-то степени на них могут влиять различные культурные традиции. Даже если мы по отношению к кому-либо проявляем жестокость, мы вполне отдаем себе отчет, что заставляем этого человека страдать. Мы автоматически предполагаем, что его психические переживания должны быть подобны тем, которые испытывали бы мы сами в той же ситуации. Похоже на то, что естественный отбор сделал нас такими, что мы предполагаем сходство между психическими переживаниями других людей и нашими собственными. Почему?
Некоторые исследователи (например, Humphrey, 1979; Crook, 1980) утверждают, что в ходе эволюции сообществ, члены которых были тесно связаны между собой, определенные преимущества имели те сообщества, где особи оказывались способными понимать других. Под «пониманием других» мы подразумеваем определенную осведомленность о других как о существах, чувства которых подобны нашим собственным. Такое понимание могло бы помочь развитию способности особей реагировать на индивидуальные качества других особей, а быть может, и развитию языка, способного выражать эти симпатии. Гриффин (Griffin, 1981) полагает, что то же самое могло происходить с представителями других видов животных и что «животные, которые осознают свои социобиологические цели, могут достигать их более эффективно, чем было бы в противном случае». Можно с определенностью сказать, что социальная и политическая жизнь некоторых приматов 1) кажется достаточно сложной,чтобы обеспечить такого рода развитие. Проблема заключается в том, что для каждой обнаруженной нами или предполагаемой характерной особенности живого организма мы всегда можем выдумать кажущееся правдоподобным адаптивное преимущество.
1) Мы полагаем, что в данном случае не лишенный юмора автор имеет в виду человека. - Прим. перев.
Мы можем достичь больших успехов, если сконцентрируем внимание на возможности возникновения в ходе эволюции более широко распространенных и более простых качеств психики животных. Почему, например, могло развиться у животных чувство боли? Если представить себе популяцию животных, которые обладают простыми поведенческими реакциями избегания, но лишены ощущений боли или страдания, то можно также предположить появление среди них мутанта, имеющего некоторое примитивное понятие о боли. Смог бы такой мутант успешно «завоевать» популяцию? Очевидно, что это новое качество может дать преимущества в процессе отбора только в том случае, если оно вызовет какие-то изменения в поведении животного. Трудно представить себе, каким образом естественный отбор мог бы влиять на психическое переживание, если бы оно было чем-то чисто личным и никак не проявлялось вовне. Быть может, это новое качество могло бы улучшить способность животного общаться со своими родственниками, а может, как-то помогло бы ему более эффективно узнавать об опасностях окружающего мира. Мы этого не знаем, но мы можем по крайней мере отточить наши гипотетические представления, проверяя их соответствие известным закономерностям эволюционной биологии. Смогло ли бы предполагаемое новое свойство успешно завоевать популяцию? Смогла ли бы сформироваться эволюци-онно стабильная стратегия (см. гл. 7)? И так далее.
Рассматривая вопрос о страдании у животных с эволюционной точки зрения, надо бы «использовать сомнения в пользу обвиняемых» и предположить, что животные испытывают страдания в тех ситуациях, которых они стремятся избежать. Если животные на самом деле способны страдать, то это, по всей вероятности, происходит в ответ на воздействие таких обстоятельств, которые являются для них в функциональном плане невыгодными. В общем и целом можно ожидать, что в процессе эволюции животные оказались сформированными таким образом, чтобы выбирать ситуации, приводящие к возрастанию их общей приспособленности, и избегать тех ситуаций, которые ведут к снижению приспособленности. По всей вероятности, следовательно, мы можем использовать поведение выбора у животных для того, чтобы определить, в каких условиях они лучше себя чувствуют.
В 1880 г. Герберт Спенсер (Spencer) предположил, что субъективное переживание удовольствия и боли развилось у животных для того, чтобы помочь им выбирать подходящие места обитания и условия жизни. Хотя хорошо известно, что у диких животных проявляются явные предпочтения по отношению к определенным местообитаниям (Lack, 1937; Hilden, 1965; Partridge, 1978), это еще не означает, что для выбора местообитания им необходимы субъективные ощущения. Однако если смотреть на это с точки зрения эволюции, то можно ожидать, что предпочтение животными тех или иных место-обитании обеспечивает им определенное благополучие. Что касается домашних или лабораторных линий животных, то здесь связь между условиями, которые они выбирают, и их общей приспособленностью не столь резко выражена. Тем не менее если предоставить этим животным возможность выбора, то по их поведению можно, по-видимому, получить вполне разумную эмпирическую оценку степени их благополучия в данных условиях (Dawkins, 1980).
Ценность такого подхода можно проиллюстрировать следующим примером. Дело касается попыток уточнить требования, которым должны отвечать условия клеточного содержания кур-несушек. Правительство Соединенного Королевства создало комитет, который должен был определить, насколько хорошо содержатся домашние животные в животноводческих хозяйствах интенсивного типа. В своем отчете комитет рекомендовал не использовать в клеточных батареях полы, сделанные из тонкой шестигранной проволоки. Комитет считал, что стоять на таком полу курам неудобно (Brambell, 1965). Однако когда кур подвергли проверке в тесте на предпочтение между этим типом пола и полом, сделанным по рекомендации комитета из толстой прямоугольной металлической сетки, то исследователи обнаружили, что птицы предпочитают пол из тонкой шестигранной проволоки (Hughes, Black, 1973). Как показали фотографии, сделанные с нижней стороны пола, ячейки из тонкой проволоки дают больше опоры для ног курицы.
Мэриан Доукинс в многочисленных экспериментах исследовала вопрос о том, какие условия обитания предпочитают домашние птицы. В одном из экспериментов она предоставляла курам возможность выбирать между клеткой в закрытом птичнике и открытой вольерой в саду. Когда птицы добирались по коридору экспериментальной установки до Т-образного разветвления, они видели с одной стороны вольеру в саду, а с другой - внутренность птичника с клеточными батареями. Куры имели возможность выбрать те или иные условия. Перед тем как повторить тест, птицу в течение пяти минут держали в той обстановке, которую она выбрала. Те куры, которые до экспериментов содержались в уличной вольере, выбирали вольеру уже в самых первых тестах. Другие, которые раньше жили в клетках на птицефабрике, вначале предпочитали привычную обстановку клетки (рис. 28.2). Однако по мере многократного повторения тестов они начинали предпочитать наружную вольеру. Отсюда следует, что если незадолго до очередного эксперимента куры имели хотя бы кратковременный опыт пребывания в наружной вольере, то этого было достаточно, чтобы изменить их выбор в тесте на предпочтение (Dawkins, 1976; 1977).
Тесты на предпочтение уязвимы для критики. Дело в том, что на выбор, который делают животные, влияет целый ряд различных факторов. Так, на предпочтение тех или иных условий обитания могут влиять генетические факторы, импринтинг, степень знакомства животного с предлагаемыми условиями или память о тех условиях, в которых находилось животное в последнее время. Генетически обусловленные различия можно исследовать, тестируя разные генетические линии домашних животных (Dawkins, 1980). Ранний опыт животных может оказывать долговременное влияние на предпочтение ими тех или иных условий обитания, но если исследования проводятся на домашних животных, то это влияние можно контролировать.
При проведении экспериментов, основанных на реакции выбора, необходимо убедиться, что животные знакомы с предлагаемыми им альтернативами. Многие животные избегают новых и незнакомых ситуаций. Совершенно очевидно, что они вполне могут предпочесть тот вариант условий, который им уже знаком. Фермеры иногда говорят, что их питомцам, по-видимому, нравятся условия их содержания, потому что, если выпустить их на свободу, они возвращаются обратно. Это напоминает рассуждения о том, что некоторые люди предпочитают сидеть в тюрьме, потому что после длительного заключения им трудно приспособиться к жизни на свободе. Проблема состоит в том, чтобы понять, каким образом взаимодействуют между собой степень знакомства животного с ситуацией и его жизненный опыт, полученный в самое недавнее время. В начале экспериментов выбор животного может определяться тем, в какой обстановке оно находилось в самое последнее время. Однако такое предпочтение может быть очень недолгим, как в описанном выше случае с курами, которых содержали в клеточных батареях.

Рис. 28.2. Курица в одном из экспериментов Мэриан Доукинс. Куры, которых в течение нескольких недель содержали в клеточных батареях, в тесте на предпочтение между клеткой в клеточной батарее и уличной вольерой могут вначале выбрать привычную для них клетку. (Фотография Tony Alien.)
Тесты на предпочтение подвергались критике еще и потому, что они не учитывали разницы между предпочтением, которое проявляется в течение короткого интервала времени, и стойким, долговременным предпочтением. Дункан (Duncan, 1977; 1978) показал, что куры предпочитают войти в гнездо-ловушку, чтобы снести яйца, даже несмотря на то, что в результате им приходится несколько часов просидеть взаперти без доступа к пище. Предпочтительное отношение к гнезду настолько сильно, что такое поведение повторяется изо дня в день. Однако оценить такого рода явления можно только с учетом имеющихся у животного альтернатив. Каким образом распределило бы животное свое время, если бы могло выбирать между различными «режимами» дня? Можно ожидать, что в дикой природе животные организуют свой ежедневный распорядок жизни наилучшим возможным образом (см. гл. 16). Однако перестройка с одного распорядка дня на другой может происходить медленно. У животных имеются как кратковременный, так и долговременный механизмы приспособления к изменениям окружающей среды, и их влияние на поведение оказывается достаточно сложным. Доукинс (1980; 1982) предположила, что измерение «эластичности» поведения (см. разд. 24.6) поможет преодолеть эти трудности. Можно утверждать, что предпочтение, проявляемое животным в условиях определенного давления среды, будет давать более надежные результаты, чем выбор под влиянием праздного каприза. Домашние животные обычно не испытывают тех временных и энергетических ограничений, которые воздействуют на поведение диких животных. Поэтому нам следовало бы, быть может, создать такого рода «давление обстоятельств» при проведении наших тестов на предпочтение. Экономисты знают, что явное предпочтение потребителя по отношению к каким-то товарам может резко измениться при изменении цен. Мы могли бы получить более четкое представление о системе предпочтений у животного, если бы построили какой-то эквивалент функции спроса (см. разд. 24.6).
Будь у нас такие данные, мы могли бы, исходя из эволюционных представлений, сделать следующее утверждение: если животное готово платить определенную цену за то, чтобы устранить отклонения от своего обычного образа жизни, то это хороший показатель его благополучия. А если не на основе таких отклонений, то как еще могло возникнуть страдание?
ДЛЯ ЗАПОМИНАНИЯ
1. Животные, по-видимому, осведомлены о своем соматическом состоянии и своем поведении, но для этого не обязательно необходимо сознание.
2. Физиологические проявления эмоций наблюдаются у многих животных, но это не означает, что животные переживают эмоции точно так же, как люди.
3. Пытаясь судить о том, испытывают ли животные страдания, мы вынуждены делать такие предположения об их психическом состоянии, которые не поддаются научной проверке.
4. Рассуждая с эволюционных позиций, мы приходим к выводу, что животные обычно знают, что для них хорошо, и руководствуются этим в ситуациях выбора.
Рекомендуемая литература
Dawkins M.S., 1980. Animal Suffering, London, Chapman and Hall. Galiup G. G., Jr., (1979). "Self-awareness in primates", American Scientist, 67, 417-421. . Griffin D.R., 1976, 1981. The Question of Animal Awareness, New York, The Rockfeller University Press.
(Последние исправления - 8.1.2002)