В МИРЕ НАУКИ.
Геномный импринтинг
Scientific American. Издание на русском языке). 1990. №12. стр.14-20
Даже если от обоих родителей потомки получают идентичные гены, действие этих генов может быть различным. Гены несут родительский «отпечаток», различный у самцов и самок, который влияет на нормальное развитие организма, а также играет роль в возникновении заболеваний
КАРМЕН САПИЕНЦА
В СЕРЕДИНЕ XIX в. Грегор Мендель в своих опытах по скрещиванию гороха сделал наблюдение, которое впоследствии стало настоящей аксиомой для генетиков. Он обнаружил, что, если скрестить гомозиготное растение, имеющее гладкие семена, и гомозиготное растение с морщинистыми семенами, в потомстве все растения будут одинаковыми и дадут только гладкие семена. Этот результат не зависел от того, у какого из растении, взятых для скрещивания, - мужского или женского - семена были гладкими. Так Мендель открыл принцип эквивалентности реципрокных скрещиваний: у потомства ген действует одинаково независимо от того, от кого из родителей он унаследован.
Трудно переоценить значение этого наблюдения Менделя в истории и практике генетики. Было установлено, что такой закономерности подчиняется большое число наследственных признаков - не только у гороха, но и у многих других организмов.
Однако недавно генетики и эмбриологи описали ряд признаков, которые не подчиняются данной закономерности. Эти признаки управляются так называемым геномным импринтингом, который состоит в том, что гены, передаваемые потомству, несут специфический «отпечаток» пола родителя, т. е. отцовские и материнские гены маркированы по-разному; причем эти «отпечатки» временные и могут быть «стерты». Вследствие геномного импринтинга потомки, получившие маркированные гены от матери, отличаются от тех, которые унаследовали такие гены от отца. Другими словами, в некоторых случаях важно, от которого из родителей унаследован ген.
Многие исследователи пытались установить молекулярную природу геномного импринтинга, обеспечивающие его механизмы, а также число и функции маркируемых генов. Хотя эти усилия не увенчались пока явными успехами, сделано несколько замечательных открытий, которые расширяют понимание ряда раковых и наследственных заболеваний, а также некоторых других недугов. Изучение геномного импринтинга, возможно, прольет новый свет и на те явления в наследовании признаков, которые вполне удовлетворительно объясняются в рамках классической менделевской генетики.
Исключения из правила идентичности гибридов при реципрокных скрещиваниях (т. е. скрещиваниях между двумя формами, когда каждая из них в одном случае берется в качестве матери, а в другом - в качестве отца) в действительности известны давно, однако, как правило, их можно было отнести к одному из двух классов. Первый составляют признаки, которые определяются генами, расположенными в половых хромосомах: у самок млекопитающих в ядрах клеток имеется по две Х-хромосомы, у самцов - по одной Х- и одной У-хромосоме. Например, цветовая слепота и гемофилия связаны с генами X-хромосомы. Наследование этих сцепленных с полом признаков подчиняется вполне определенным правилам, согласно которым гибриды в реципрокных скрещиваниях не обязательно идентичны. Скажем, у отца-дальтоника и нормальной по этому признаку матери ни один из сыновей не будет дальтоником. Если мать страдает дальтонизмом, а отец нет, то все сыновья окажутся дальтониками. В обоих случаях дочери будут нести ген, обусловливающий дальтонизм, но иметь нормальное зрение. Наследование и проявление признаков, сцепленных с полом, зависят от пола потомка, но не связаны непосредственно с полом того родителя, от которого унаследован признак.
Второй класс неэквивалентных реципрокных скрещиваний включает признаки, определяемые внеядерными генами. Некоторые клеточные органеллы - митохондрии в клетках животных, митохондрии и хлоропласты в клетках растений - обладают своей собственной генетической информацией. Эти органеллы передаются из поколения в поколение с цитоплазмой яйцеклеток и поэтому наследуются исключительно по материнской линии. Таков характер наследования цвета листьев у некоторых растений, а также заболевания человека, известного под названием митохондриальной энцефаломиопатии.
Но есть еще одна группа исключений из правила эквивалентности реципрокных скрещиваний, которая разительно отличается от двух, описанных выше - это геномный импринтинг. Подверженные его действию признаки вовсе не обязательно определяются генами половых хромосом (хотя и это бывает) и совершенно не связаны с клеточными органеллами, наследующимися по материнской линии. Геномный импринтинг - по меньшей мере теоретически - может затрагивать любой ген. В основе исключений, связанных со сцеплением с полом или с наследованием органелл по материнской линии, лежат различия в генетическом вкладе родителей в генотип потомков. В случае геномного импринтинга, напротив, оба родителя могут передавать потомкам совершенно идентичные гены, но при разном - материнском либо отцовском - импринтинге действие этих генов будет неодинаковым. Эти необычные свойства привлекли внимание биологов.
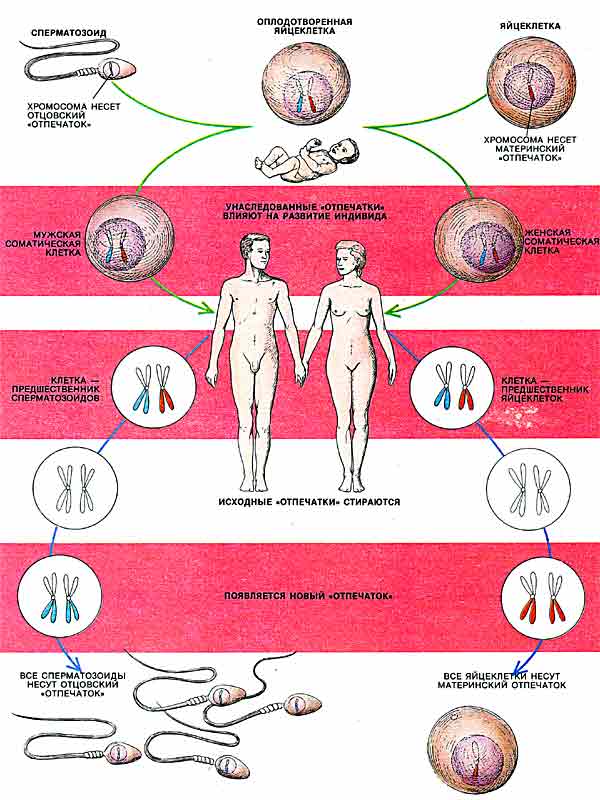
ГЕНОМНЫЙ ИМПРИНТИНГ состоит в том, что хромосомы половых клеток (сперматозоидов или яйцеклеток) индивида приобретают «отпечаток» его пола. Потомство получает один набор хромосом с отцовской маркировкой некоторых генов, а другой - с материнской. При образовании у потомка половых клеток прежний «отпечаток» стирается и эти гены маркируются в соответствии с полом данной особи.
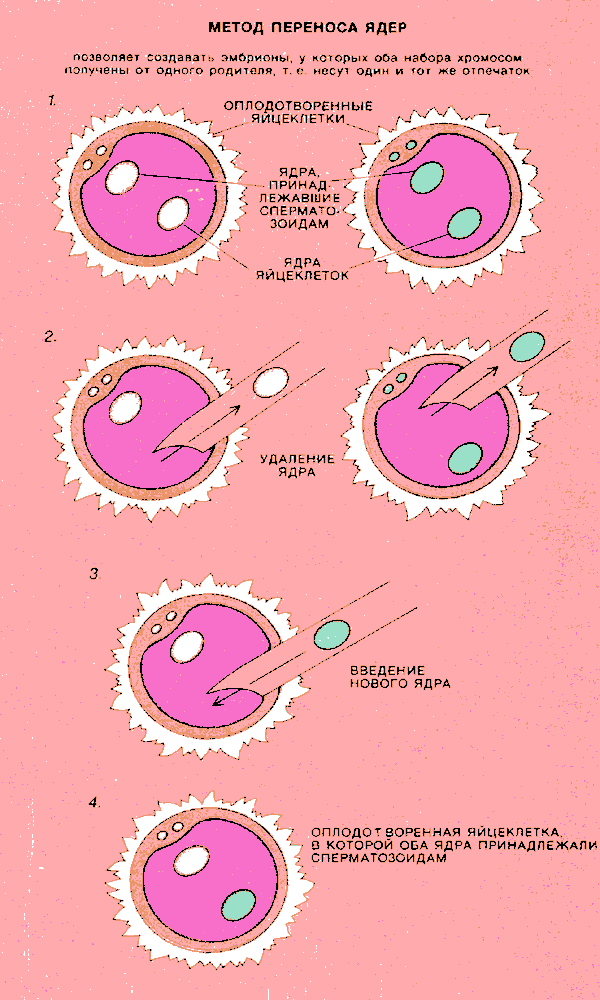
ИНТЕРЕС к признакам, подверженным действию геномного импринтинга, был вызван главным образом опытами Дж.Макграта и Д.Солтера из Вистаровского института анатомии и биологии в Филадельфии, а также М.Сурани и его коллег из Института физиологии животных в Кембридже (Великобритания). Макграт и Солтер разработали изящный микрохирургический метод переноса клеточных ядер мышиных эмбрионов, который позволил физически заменять генетическую информацию одной особи на аналогичную информацию от другой особи. Этот метод основан на том, что после оплодотворения яйцеклетки сперматозоидом зигота некоторое время не делится и ядра яйцеклетки и сперматозоида существуют в ее цитоплазме автономно друг от друга. Оба ядра хорошо видны в световом микроскопе, для них характерны определенные размеры и локализация.
С помощью тончайших стеклянных капилляров из оплодотворенной яйцеклетки избирательно удаляли ядро либо яйцеклетки, либо сперматозоида (либо оба). Затем вместо удаленного ядра в клетку вводили ядро из другой оплодотворенной яйцеклетки. Было установлено, что если заменяется ядро яйцеклетки на ядро другой яйцеклетки или же ядро сперматозоида — на ядро другого сперматозоида, то эмбрионы развиваются полностью и выросшие из них особи неотличимы от нормальных мышей той же линии.
Метод переноса дает возможность создавать необычные по генетической конституции эмбрионы, у которых оба набора хромосом унаследованы от одной родительской особи. Макграт, Солтер и Сурани получили гиногенетических (т. е. имеющих два полных набора материнских хромосом) и андрогенетических (т. е. имеющих два набора отцовских хромосом) эмбрионов и сравнили их развитие с развитием нормальных эмбрионов.
Важно отметить, что в этих экспериментах использовались инбредные линии, поддерживавшиеся в течение многих поколений, так что самцы и самки обладали идентичными наборами хромосом (за исключением, конечно, Х- и У-хромосом). Если развитие эмбриона зависит только от набора его генов, то теоретически не имеет значения, что особь получает все гены от одного родителя, а не от двух, т. е. гиногенетические, андрогенетические и нормальные мыши должны развиваться одинаково.
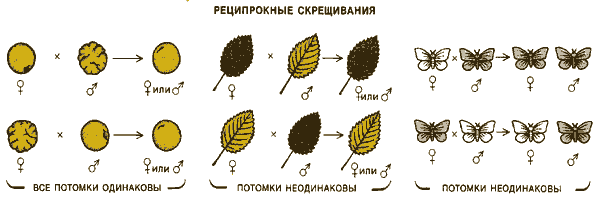
РЕЦИПРОКНЫЕ СКРЕЩИВАНИЯ выявляют, зависит ли данный признак от того, кто из родителей - мать или отец - передал потомству ген, определяющий этот признак. Для большинства признаков, например для гладкости семян у гороха, не важно, от которого из родителей получен соответствующий ген (слева). Некоторые признаки, например цвет листьев у ряда растений, наследуются исключительно от матери (в середине). Для признаков, сцепленных с полом, например окраски крыльев у пядениц Abraxas, картина наследования более сложная (справа).
В действительности эти три группы животных развивались совсем не одинаково. Эмбрионы с двумя наборами хромосом от одной и той же родительской особи — самца или самки — не развиваются до конца. Обычно их развитие останавливается на ранней стадии, когда тело эмбриона состоит всего из нескольких десятков клеток. Если же развитие все-таки продолжается, что иногда случается, то у гиногенетических и андрогенетических особей имеют место поразительно различные аномалии. У гиногенетических эмбрионов, достигших наибольшего развития, в теле эмбриона наблюдались незначительные аномалии, однако их плаценты и желточные мешки (эти структуры необходимы для питания эмбриона) оказались сильно недоразвитыми. А у наиболее развитых андрогенетических эмбрионов, наоборот, желточные мешки и плаценты были почти нормальными, а тела — мелкими и слабо развитыми.
Поскольку у гиногенетических, андрогенетических и нормальных эмбрионов последовательности нуклеотидов хромосомной ДНК не различались, Макграт, Солтер и Сурани заключили, что гены были каким-то образом модифицированы и несли «отпечаток» пола той особи, от которой они унаследованы. По-видимому, в результате этого геномного импринтинга избирательно инактивируются определенные гены, которые в норме функционируют в ранний период эмбрионального развития. Среди генов, унаследованных от отца, не проявляли активности некоторые гены, управляющие эмбриональным развитием; гены, важные для образования желточного мешка и плаценты, оказывались неактивными, если были получены от матери.
Геномный импринтинг был наглядно продемонстрирован также Б. Кэттанаком из Совета по медицинским исследованиям в Оксфорде. В течение многих лет Кэттанак изучал неэквивалентные реципрокные скрещивания. Для этих исследований он вывел линии мышей, у которых некоторые хромосомы попарно сцеплены. Поскольку у таких животных некоторые хромосомы физически соединены, они не способны расходиться в процессе клеточного деления, называемого мейозом (при котором образуются половые клетки — яйцеклетки и сперматозоиды), так что одна из клеток, возникающих в результате деления, получает обе парных хромосомы, а другая ни одной.
Если яйцеклетка с двумя хромосомами одной из пар оплодотворяется сперматозоидом, лишенным такой хромосомы, то у эмбриона число хромосом будет нормальным, однако обе хромосомы этой пары будут материнского происхождения. Точно так же обе парных хромосомы будут отцовскими, если сперматозоид нес две хромосомы данной пары, а яйцеклетка ни одной. Согласно правилу эквивалентности в реципрокных скрещиваниях, в потомстве все особи должны быть идентичными. Но из опытов Кэттанака однозначно явствовало, что для некоторых хромосом у мыши это правило не выполняется.
Например, Кэттанак обнаружил, что мыши, унаследовавшие обе хромосомы 11 от одного и того же родителя, разительно отличаются от нормальных по весу и размеру, хотя выращивались в одинаковых условиях. Мышата с двумя материнскими хромосомами 11 были ненормально мелкими, а с двумя отцовскими хромосомами 11 необыкновенно крупными. Подобные эксперименты с сцепленными хромосомами других пар выявили иные характерные нарушения, например смерть в эмбриональном возрасте и отклонения в поведении.
Кэттанак также показал, что в следующем поколении эффект геномного импринтинга не сохраняется. Значит, «отпечаток» — это непостоянная модификация хромосомы. У мелкого самца, получившего обе хромосомы 11 от матери, потомство бывает обычно нормального размера. Следовательно, материнские гены должны каким-то образом освобождаться от «отпечатка» самки и маркироваться заново как гены самца.
Воодушевленные открытиями Макграта, Солтера, Сурани и Кэттанака исследователи в нескольких лабораториях (в том числе и в лаборатории автора) пытались установить, не является ли механизмом геномного импринтинга непосредственная модификация ДНК. В частности, мы изучали роль метилирования ДНК, состоящего в том, что к молекуле ДНК химически присоединяются метильные группы СН3 (см. статью: Р. Холлидей. Эпигенетическая наследственность, «В мире науки», 1989, № 8). Действительно, эксперименты во многих случаях показали, что определенные гены метилированы по-разному в зависимости от того, получены они от матери или от отца, а также что эти модификации ДНК могут исчезать. Однако до сих пор не удалось доказать, являются ли такие модификации главным механизмом геномного импринтинга или лишь отражением какого-то пока неизвестного более общего биологического явления.
Не все мои коллеги согласятся со мной, но я готов держать пари, что метилирование ДНК — это только следствие основного механизма импринтинга. К счастью, можно ставить важные эксперименты, не зная действительных причин явления. Каков бы ни был механизм геномного импринтинга, его проявления часто странны и поразительны.
ПРОЯВЛЕНИЯ геномного импринтинга удивительным образом связаны с некоторыми заболеваниями человека. Неожиданно оказалось, что в природе уже существуют параллели тем экспериментальным состояниям, которые Кэттанак исследовал у мышей. Недавно Р. Николлс и его коллеги из Медицинской школы Гарвардского университета установили, что у многих больных с синдромов Прадера—Вилли (при этом заболевании характерны умственная отсталость, крайнее ожирение, низкий рост и непропорционально малый размер рук и ног) обе хромосомы 15 унаследованы от матери.
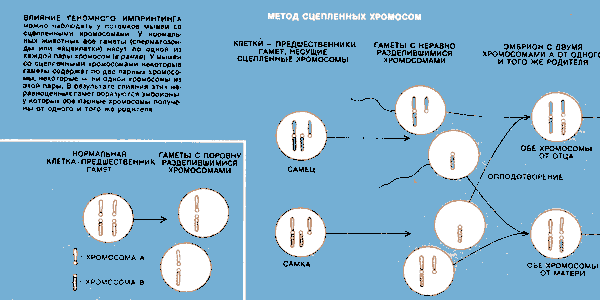
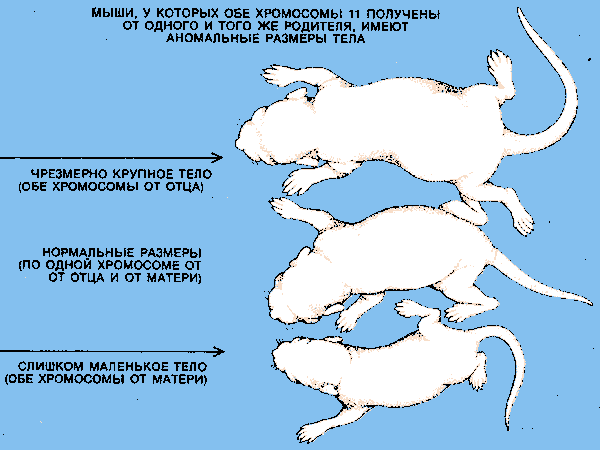
ВЛИЯНИЕ ГЕНОМНОГО ИМПРЙМТИНГА можно наблюдать у потомков мышей со сцепленными хромосомами. У нормальных животных все гаметы (сперматозоиды или яйцеклетки) несут по одной из каждой пары хромосом (в рамке). У мышей со сцепленными, хромосомами некоторые гаметы содержат no две парных хромосомы, некоторые — ни одной хромосомы из этой пары, В результате слияния этих неравноценных гамет образуются эмбрионы, у которых обе парные хромосомы получены от одного и того же родителя
Р. Николлс, Дж. Кнолл (тоже сотрудник Гарвардского университета) и Ч. Уильямс из Флоридского университета обнаружили связанную с геномным импринтингом закономерность у больных с синдромом Ангельмана, при котором наблюдается неадекватная смешливость, резкие судорожные движения и другие симптомы двигательной и умственной отсталости. У таких больных нередко имеют место частичные делеции унаследованной от матери хромосомы 15, в результате чего полностью функциональна только отцовская хромосома 15.
Эти два заболевания, хотя и столь разные по клинической картине, могут быть связаны с различиями в импринтинге одних и тех же генов одной и той же хромосомы. Однако в отличие от ненормально крупных и мелких мышей в экспериментах Кэттанака, синдромы Прадера—Вилли и Ангельмана не удается представить просто как две стороны одной медали, т. е. объяснить избытком либо недостатком продуктов одних и тех же генов. Исследования Николлса и его коллег показали, что бывает очень трудно предсказать, каким образом конкретные признаки зависят от процесса геномного импринтинга. Полезно было бы изучить на этот предмет многие наследственные заболевания человека; не исключено, что найдутся указания на влияние геномного импринтинга.
Геномный импринтинг может играть роль даже в тех состояниях, которые удается объяснить без влияния этого явления. Для иллюстрации рассмотрим несколько примеров детского рака — эмбриональную рабдомиосаркому (рак мышц), опухоль Вильмса (рак почек) и остеосаркому (рак костей).
Чтобы показать, каким образом геномный импринтинг может участвовать в развитии этих заболеваний, напомним представления о возникновении рака. Считается, что многие виды рака являются результатом накопления мутаций в определенных генах одной и той же клетки. Например, хромосома 11 несет ген, известный под названием Rd, который относится к семейству генов, называемых рецессивными онкогенами, генами-супрессорами рака или антионкогенами (см. статью: Р. Вайнберг. Поиск антионкогенов, «В мире науки», 1988, № 11). Лишенная продукта гена Rd, мышечная клетка претерпевает злокачественную трансформацию и дает начало рабдомиосаркоме.
Поскольку в ядре каждой клетки имеются две хромосомы 11, для злокачественной трансформации необходимо, чтобы инактивировались обе копии гена Rd. Первая из них может быть выведена из строя разными путями; самый простой — это мутация, изменяющая последовательность нуклеотидов ДНК в самом гене. Если такая мутация произошла, то любое дополнительное генетическое событие, в результате которого инактивируется оставшаяся копия Rd, вероятно, приведет к злокачественной трансформации клетки. Нередко это событие заключается в утрате целой хромосомы или в делении ее части, содержащей данный ген.
Мои коллеги X. Скрэбл и У. Кавени из Онкологического института Людвига в Монреале и я предложили альтернативный механизм возникновения некоторых видов рака у детей. Мы предполагаем, что первая копия рецессивного онкогена инактивируется не истинной мутацией, а из-за геномного импринтинга, при котором избирательная инактивация генов происходит по-разному у мужчин и у женщин. Вторым событием, которое инициирует злокачественную трансформацию клетки, является скорее всего утрата хромосомы или ее части. Согласно нашей гипотезе, в клетках рабдомиосаркомы в сохранившейся хромосоме 11 ген Rd должет быть инактивирован в результате импринтинга, причем в большинстве случаев эмбриональной рабдомиосаркомы обе хромосомы 11 должны наследоваться от одного и того же родителя.
В нашей лаборатории, а также в лабораториях Г. Сондерса из Онкологического центра М. Д. Андерсона в Хьюстоне, М. Манненса из Амстердамского университета, М. Сазаки из Университета в Киото и ряде других получены данные, свидетельствующие в пользу этой гипотезы. Выяснилось, что при эмбриональной рабдомиосаркоме, опухоли Вильмса и остеосаркоме первая инактивация почти всегда затрагивает рецессивный онкоген, находящийся в отцовской хромосоме. Такие данные подтверждают участие геномного импринтинга в развитии упомянутых видов рака, но они же создают серьезные теоретические затруднения.
ТРУДНОСТИ возникают, поскольку геномный импринтинг рассматривался как следствие огромных физиологических и биохимических различий в образовании гамет (сперматозоидов и яйцеклеток) у самцов и самок. На протяжении почти всего периода половой зрелости у самцов из постоянно делящихся стволовых клеток образуются миллиарды сперматозоидов. Самки, напротив, от рождения обладают всеми яйцеклетками, которые будут созревать в организме на протяжении репродуктивного периода жизни. По мере того как эти яйцеклетки созревают, одна или несколько одновременно, происходит регулярная овуляция, т. е. выход яйцеклеток в половые пути, где происходит оплодотворение.
Если бы различия в геномном импринтинге были лишь отражением разных путей образования гамет у самцов и у самок, то, вероятно, у всех самцов он происходил бы одинаково, а у самок по-другому, но тоже одинаково у всех. За отсутствием данных, которые бы говорили об обратном, мы предположили, что у животных одного пола геномный импринтинг осуществляется одинаково.
Однако данные о связи рака в детском возрасте с неактивными рецессивными онкогенами отцовского происхождения опровергли наше предположение. Если, например, у всех мужчин ген Rd подвергается импринтингу и инактивируется, то заболеваемость эмбриональной рабдомиосаркомой должна быть очень высокой, поскольку тогда любое генетическое изменение в гене Rd, унаследованном от матери, приводило бы к появлению в организме раковых клеток. Но поскольку это заболевание встречается редко (им страдает приблизительно один из 20 тыс. детей), маловероятно, что каждый человек наследует от отца неактивный рецессивный онкоген. Однако же у заболевших, по-видимому, именно это имеет место.
Это противоречие можно объяснить тем, что не у всех самцов (и, наверное, не у всех самок) одни и те же гены подвергаются импринтингу и инактивируются. Чем могут быть обусловлены подобные различия между индивидами?
Генетик всегда начинает поиски ответа на такие вопросы с генетического подхода. С этой точки зрения предполагается, что существуют один или несколько генов, ответственных за импринтинг. (Не путать эти гены с генами, которые сами подвергаются геномному импринтингу!) Другими словами, импринтинг рассматривается как процесс, определяемый нестрого фиксированной группой генов, которые у самцов и у самок работают по-разному. Поэтому у особей одного пола, но обладающих разными генами, управляющими импринтингом, должны различаться и наборы генов, которые модифицируются в этом процессе. Например, у большинства мужчин ген Rd не инактивируется, но в редких случаях у индивидов, которые обладают аберрантным геном, управляющим импринтингом, инактивация гена Rd будет происходить.
Идея о том, что экспрессия гена или ее отсутствие могут определяться другими генами, не нова. По сути она описывает феномен, давно известный в генетике под названием модификации доминантности: многие признаки зависят от активности генов, которые влияют на экспрессию генов, непосредственно определяющих эти признаки. Геномный импринтинг можно считать особым случаем модификации доминантности. Для генов-модификаторов, которые управляют геномным импринтингом, постулируется лишь одно необычное свойство: у самцов и самок они должны работать по-разному.
Интересный пример признака, который модифицируется зависимым от пола геномным импринтингом, представляет болезнь Хантингтона. Это смертельное неврологическое расстройство наследуется как доминантный признак, т. е. оно проявляется у каждого человека, унаследовавшего ген, обусловливающий заболевание, хотя бы от одного из родителей. Обычно болезнь Хантингтона поражает людей немолодых; в среднем диагноз ставится в возрасте 38 лет. Однако приблизительно в 10% случаев болезнь развивается еще до достижения зрелости и бывает, что она начинается у ребенка уже в два с половиной года.
Ранее было установлено, что примерно 90% больных детей получают ген, обусловливающий заболевание, от отца. Ясно, что в этих случаях и дети, и их отцы несут один и тот же ген, ответственный за развитие болезни; однако его модификация в результате отцовского импринтинга приводит к тому, что заболевание проявляется в значительно более раннем возрасте.
Для объяснения генетики болезни Хантингтона выдвигались разнообразные гипотезы, однако лучше всего имеющиеся данные согласуются с моделью, предложенной Ч.Лэйрдом из Вашингтонского университета. Нововведение Лэйрда заключается в том, что он провел аналогию с особым классом признаков дрозофилы, характеризующихся изменчивой экспрессией, или мозаичным эффектом положения. Необычность этих признаков в том, что в отличие от большинства других признаков, обусловленных мутациями, они проявляются не во всех клетках ткани. Ткани в таких случаях оказываются как бы мозаичными: часть клеток в них имеет нормальный, а часть мутантный фенотип. Доля клеток, в которых экспрессируется мутация, определяется действием генов-модификаторов, в зависимости от которого баланс мозаицизма может сдвигаться от почти полностью нормальных до почти полностью мутантных тканей.
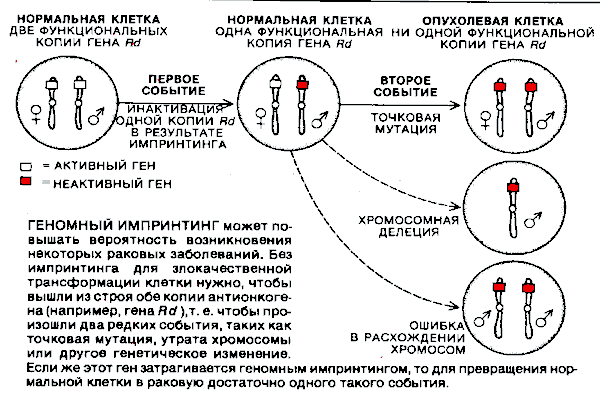
ГЕНОМНЫЙ ИМПРИНТИНГ может повышать вероятность возникновения некоторых раковых заболеваний. Без импринтинга для злокачественной трансформации клетки нужно, чтобы вышли из строя обе копии антионкогена (например, гена Rd ),т. е. чтобы произошли два редких события, таких как точковая мутация, утрата хромосомы или другое генетическое изменение.
Если же этот ген затрагивается геномным импринтингом, то для превращения нормальной клетки в раковую достаточно одного такого события.
Лэйрд предположил, что если различия в возрасте начала болезни Хантингтона отражают мозаицизм в экспрессии гена, обусловливающего заболевание, то этот мозаицизм, вероятно, зависит от генов-модификаторов. Если ген-модификатор действует так, что баланс мозаицизма сдвигается в сторону мутантных клеток и возникает почти полностью мутантная ткань, то заболевание развивается рано; при противоположном действии гена оно проявляется в позднем возрасте. (Описаны случаи, когда болезнь Хантингтона начиналась у стариков на седьмом десятке.)
Тот факт, что при наследовании гена, обусловливающего заболевание, от отца, в большинстве случаев болезнь начинается в раннем возрасте, Лэйрд объяснил расположением гена-модификатора в Х-хромосоме. Поскольку мужчины имеют по одной X-хромосоме, любая аномалия гена-модификатора будет проявляться, а не компенсироваться, как у женщин, имеющих по две Х-хромосомы. Поэтому для мужчины больше вероятность того, что у его потомков баланс мозаицизма окажется сдвинутым в сторону большей доли мутантных клеток в тканях и болезнь разовьется в раннем возрасте. В 10% случаев раннего развития болезни Хантингтона ребенок получает соответствующий ген от матери, а не от отца; этого и следует ожидать, так как хотя и редко, но должны встречаться женщины, у которых ген-модификатор аномален в обеих Х-хромосомах.
Судя по всему, модель Лэйрда хорошо описывает генетику болезни Хантингтона, которой свойственна сложная картина проявления. В своей работе я слегка изменил эту модель, попытавшись конкретизировать, каким образом действуют гены-модификаторы, что позволяет точнее объяснить наблюдаемые генетические особенности данного заболевания.
На мой взгляд, модель Лэйрда, объясняющая генетику болезни Хантингтона мозаичным эффектом положения, замыкает некий цикл исследований геномного импринтинга. Первым глубоким исследованием геномного импринтинга является работа Дж. Споффорд из Чикагского университета, изучавшей мозаичный эффект положения у дрозофилы. Статья Споффорд была опубликована в 1959 г., но не привлекла должного внимания. За последние 30 лет время от времени появлялись и другие сообщения о наследственных признаках, зависящих от пола родителя, от которого унаследован соответствующий ген. По-видимому, геномный импринтинг влияет на признаки у самых разных организмов - дрозофилы, дрожжей, кукурузы, мыши и человека.
Тем не менее геномный импринтинг принято считать генетическим курьезом, затрагивающим лишь очень немногие признаки. Меня неоднократно спрашивали, почему я попусту трачу свое время (и, как подразумевалось, время вопрошающего) на столь незначительное явление. Всякий раз я отвечаю, что число признаков, подверженных влиянию геномного импринтинга, не известно, но, вероятно, очень велико. Обычно такой ответ вызывает удивленно-скептический взгляд и коротенькую лекцию о законах Менделя, завершающуюся безапелляционным утверждением, что к большинству признаков геномный импринтинг не имеет никакого отношения.
До какой-то степени эти критики правы. Если бы для большинства признаков существовала четкая зависимость их проявления от пола родителя, от которого унаследован соответствующий ген, генетики, несомненно, заметили бы это. Однако далеко не все признаки изучены одинаково подробно. Когда Споффорд исследовала мозаичную экспрессию мутантных признаков у дрозофилы, она, как и Мендель, наблюдала одни и те же признаки у потомства в реципрокных скрещиваниях. И все-таки различия в экспрессии признаков оказались достаточными, чтобы она подметила зависимость экспрессии от пола родителя, несущего мутацию.
Точно так же работы, проведенные в моей лаборатории, а также в лаборатории моего коллеги А.Петерсона из Онкологического института Людвига, показали, что некоторые признаки у мышей являются мозаичными. Если бы в этих опытах задача заключалась лишь в том, чтобы установить, присутствуют ли данные признаки, то она решалась бы однозначно: да, присутствуют; однако, если выяснять, какова экспрессия этих признаков, то обнаруживается влияние геномного импринтинга. Для одних признаков степень экспрессии может быть и не важна, но для других она оказывается решающей, например определяет возникновение опухоли Вильмса или возраст проявления болезни Хантингтона.
Одной из наиболее многообещающих областей исследования геномного импринтинга, вероятно, является изучение наследования сложно проявляющихся болезней человека. Насколько плодотворен такой подход, покажет время, однако успех - если и не практический, то теоретический, - которого мы достигли в понимании болезни Хантингтона и некоторых видов детского рака, наполняет меня уверенностью, что прогресс будет достигнут.
(Последние исправления - 28.5.2002)