
Глава 5. ОБЩЕСТВЕННОЕ ПОВЕДЕНИЕ ЖИВОТНЫХ
Общественное поведение — одна из важнейших форм видоспецифического поведения, изучение которого составляет обширный раздел этологии с момента ее возникновения и до настоящего времени. Благодаря многочисленным исследованиям типы организации сообществ (или социодемографических систем) и характер внутригрупповых отношений описаны достаточно подробно у большого числа видов. Наибольшее внимание при этом было уделено структуре сообществ у млекопитающих и птиц, однако мы рассмотрим также и такое замечательное явление, как общественное поведение насекомых (см.: 5.6).
Исследование закономерностей существования и развития таких объединений у многих видов животных способствует пониманию этологической структуры популяций. Эти вопросы подробно проанализированы в монографии Е.Н.Панова (1983).
Характер внутривидовых взаимоотношений особей тесно связан с различными типами их распределения в пространстве. Как правило, исследователи выделяют два основных типа использования территории. Первый — это не перекрывающиеся между собой и не защищаемые участки обитания отдельных особей или семей, второй — не перекрывающиеся, но защищаемые территории. Первый тип характерен для "социальных" видов животных, второй — для "территориальных" или одиночных видов (см.: 5.2.1). Даже у животных, относящихся к одиночным видам, существуют периоды жизненного цикла, когда они вступают в контакт с особями своего вида, например в сезон размножения. Взаимоотношения между самцами и самками, в особенности в брачный период, также относятся к категории внутривидовых общественных отношений.
Минимальная социальная ячейка, цельность и самостоятельность которой поддерживаются в основном ее собственными, внутренними механизмами, называется микропопуляцией. Феномен микропопуляции особенно подробно изучался у домовой мыши, и применительно к этому виду микропопуляция называется "демом". По определению Н.П. Наумова (1977), подобное объединение особей именуется "парцеллой".
Исследование динамики численности популяций показало важную роль поведения особей в этих процессах (Шилов, 1977).
Видоспецифические особенности социальной организации более заметны в процессе формирования группы, а не в стабильный период ее существования, В качестве примера в разделе 5,5.1 сравниваются социальные отношения и особенности агрессивных контшстов у нескольких близкородственных видов песчанок, Оригинальный научный подход, реализованный в этом исследовании (Гольцман и др., 1983; 1994), представляет собой, с одной стороны, прямое развитие классических этологических методов (наблюдение в естественной или "полуестественной" среде, сравнение близкородственных видов), а с другой — их принципиально новое развитое, поскольку он позволяет перенести внимание со статичной картины структуры группы на динамику ее формирования.
В ранний период развития науки о поведении описание сообществ животных ограничивалось, как правило, констатацией фактов существования внутригрупповых отношений и их характеристикой.
Одна из первых работ, в которой был проведен анализ особенностей поведения отдельных особей в сообществе и выявлена иерархическая структура в группах домашних кур, была опубликована Т. Шьелдерупп-Эббе в 1924 г. (подробнее об иерархии доминирования см.: 5.3.1).
Важным этапом в изучении общественного поведения животных стали две знаменитые работы К. Лоренца: "О поведении общественных видов врановых птиц" (Lorenz, 1927) и "Компаньон в мире птиц" (Lorenz, 1935). В них Лоренц описал свои многолетние наблюдения за колонией индивидуально распознаваемых (благодаря цветным меткам) галок и большой стаей приручаемых из поколения в поколение диких гусей. На основании этих наблюдений Лоренц делает заключение, что и стая гусей, и колония галок — это не случайные группировки отдельных особей. Это организованные сообщества с четкой внутренней структурой и сложными взаимоотношениями ее членов (см. также: Лоренц, 1978; 1994).
К.Лоренц провел анализ возможных эволюционных корней такого важнейшего феномена социального поведения, как "ритуалы" — одного из основных способов общения животных (см.: 5.3.3). Было установлено, что поддержание лидирующего положения в группах многих видов животных осуществляется не за счет драк и нападений, а преимущественно за счет ритуалов, т.е. обмена соответствующими сигналами и позами, которые называют демонстрациями. Как правило, такие ритуалы представляют собой фиксированные комплексы действий (ФКД, см.: 3.3.2), и их значение бывает понятно всем членам данного вида без специального обучения. Анализируя этот вид поведения птиц, Лоренц продемонстрировал возможность возникновения ритуалов в процессе эволюции путем постепенной замены прямых нападений угрозами и запугиванием соперника, К. Лоренц и его коллега смогли показать, что так называемая "триумфальная церемония" имеет важнейшее значение в общении серых гусей (рис. 5.1). Это характерное поведение выполняется после нападения одного из членов группы на действительного или мнимого противника. Оно подробно описано в книгах "Год серого гуся" и "Агрессия" (Лоренц, 1984; 1994).
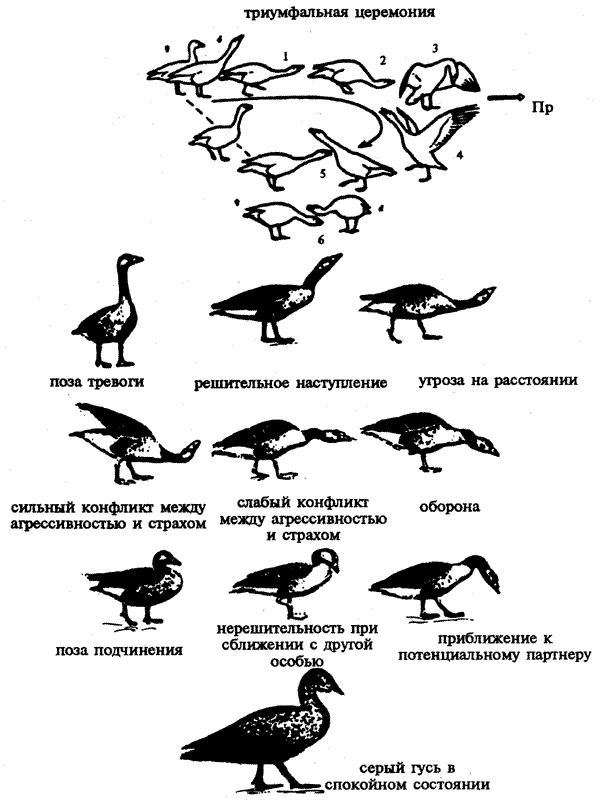
Рис.5.1. Этологическое описание поведения серого гуся. Вверху — последовательность смены поз при триумфальной церемонии. Самец надвигается (1 и 2) на противника (Пр), реального или воображаемого, и его прогоняет (3), затем разворачивается к самке, принимая определенную позу и издавая крик. Самка движется ему навстречу, также издавая крик. Затем они, находясь рядом, кричат и гогочут (5 и 6). Движения шеи гуся при угрозе и приветствии сходны, что делает очевидным происхождение этого брачного ритуала от агрессивных демонстраций. Ниже приводятся — отдельные позы серого гуся, имеющие то или иное "значение" (по Лоренцу)
Было показано, что отдельные компоненты данного ФКД отражают разные оттенки взаимоотношений между птицами. Эти работы вместе с исследованиями Н.Тинбергена положили начало изучению основ коммуникации, или языка животных (см.: 5.4). В работах Н.Тинбергена и его коллег сопоставлялось поведение, в том числе ритуалы и демонстрации, у близкородственных видов чаек (см.: 5.3.3). В частности, была исследована роль конфликта противоположных стремлений (агрессивности и страха, агрессивности и полового побуждения) в появлении некоторых демонстраций (см.: 4.3). Впоследствии этот же подход был успешно использован для анализа коммуникационного поведения у животных других групп. Исследования Н. Тинбергена по эволюции инстинктивных действий и их адаптивной ценности (см.: 6.3) стали основополагающими для анализа социальной организации животных разных видов с учетом тех преимуществ, которые она дает для выживания вида по сравнению с одиночным образом жизни.
Сообществом называется такой тип внутрипопуляционных отношений, при котором особи образуют стабильные группировки, занимающие и защищающие определенную территорию. Они поддерживают постоянный обмен информацией, находятся в некоторых относительно постоянных отношениях и скрещиваются преимущественно друг с другом. Сообщество включает в себя, как правило, небольшое число особей. Это более мелкая, чем популяция, группа особей данного вида, которую иногда называют микропопуляцией. Для сравнительного изучения структурных и иных особенностей сообществ животных исследователям необходимо выделить признаки, по которым группировки особей разных видов могут различаться между собой. Важнейшими признаками при этом оказываются:
1) устойчивость группировок во времени;
2) взаимная координация действий особей в группе;
3) прочность связей между особями;
4) поддержание целостности группы (агрессия по отношению к "чужакам" своего вида).
Одиночно живущими называют животных, у которых контакты между особями поддерживаются преимущественно в период спаривания, а также (у некоторых видов) в период ухаживания за потомством. Наиболее характерными и достаточно известными примерами животных, живущих поодиночке, являются представители семейства кошачьих. Одиночный образ жизни можно считать устойчивой характеристикой большинства видов этой группы, за исключением львов. Следует отметить, что практически всегда он связан с поддержанием более или менее четко ограниченной территории, индивидуального участка, который служит местом пребывания особи и ее охотничьими угодьями. У многих крупных хищных млекопитающих (и птиц) размеры индивидуальных участков очень велики, поскольку они должны обеспечивать возобновляемые запасы дичи. Примером самого крупного одиночно живущего хищника средней полосы является бурый медведь (см.: 5.5.2).
В социобиологических построениях (см.: 6.5.1) одиночный образ жизни рассматривается как исходный в эволюции социальных структур разной сложности (см.: Панов, 1983). Эволюцию усложнения социальной структуры, как правило, изучают на примере перепончатокрылых — группы насекомых, среди которых можно найти практически все переходы от одиночных к социально организованным формам.
Примеры переходов от сугубо одиночного образа жизни к разным формам более или менее длительных семейных объединений у млекопитающих дают грызуны. У некоторых видов млекопитающих, например у волка, одиночный образ жизни составляет обязательную фазу годового жизненного цикла (см.: 5.5.2).
Уже в ранний период развития этологии был обнаружен широкий диапазон различных типов социальной организации животных, начиная от одиночного образа жизни и кончая очень сложными сообществами приматов и общественных насекомых. Сообщества, даже самые примитивные, следует отличать от скоплений, или агрегаций, животных. Это такие объединения животных, которые формируются под действием какого-то физического фактора среды (пищи, температуры и т.п.). Примером агрегации могут служить стайки головастиков в прогретых солнцем местах водоема. Скопления характерны для многих видов беспозвоночных (см.: 5.6.1)
В книге "Агрессия", вышедшей впервые в 1966 г., К. Лоренц, рассматривая различные формы социальных структур, неявно проводит следующую классификацию: все сообщества животных можно разделить на два коренным образом различающиеся класса: анонимные, в которых нет ничего похожего на структуру (ни группировок, ни вожаков, ни ведомых), и персонифицированные, основанные на личных контактах, в которых возможно распределение ролей. Этот признак — наличие или отсутствие способности членов группы узнавать друг друга — используется в качестве одного из основных критериев классификации сообществ.
Сообщество животных, персонально не знающих друг друга, называют анонимным. Было принято считать, что анонимные сообщества характерны главным образом для более низкоорганизованных групп животных, а с усложнением нервной системы и поведения в целом происходит и усложнение социальной организации. Однако это не совсем так. К таким анонимным сообществам относятся перелетные стаи, а также скопления многих видов птиц на ночевках, хотя последние могут состоять из более мелких групп, члены которых персонально знают друг друга.
Следует отметить, что и в стае перелетных птиц (анонимном сообществе), например лебедей, диких гусей и журавлей, семейные группы (супружеские пары с детьми) держатся вместе и сохраняют личные связи. Но так бывает не у всех видов. В этой же ситуации аисты и цапли — члены супружеских пар — друг друга не узнают. Когда приходит время вить гнездо, супруги действуют независимо один от другого, даже если пара сохранилась: просто самец и самка каждый сам по себе прилетают на старое место.
Анонимное сообщество часто демонстрирует удивительную сплоченность и целесообразность групповых действий. Так, стаи скворцов при появлении в воздухе ястреба-перепелятника или чеглока плотно стягиваются, спешат ему навстречу и, обтекая со всех сторон, вновь собираются у него в хвосте. Так же реагируют на хищника и многие рыбы. Хищники же, не только крупные, но и мелкие, не нападают на жертву внутри плотного стада, а стараются отбить кого-то одного или выждать, пока наиболее пугливая и беспокойная жертва сама отделится от стада (см.: Лоренц, 1994).
В анонимных сообществах особи согласованно реагируют на биологически значимые сигналы, которые могут иметь разную модальность. В птичьих колониях это громкие крики, оповещающие о приближении хищника или просто о каких-то переменах обстановки. Сигналы другой модальности воспринимаются с помощью зрения, например беспокойные движения одной особи, которые побуждают остальных членов стаи следовать за ней. Такая реакция называется кинопсисом. Сигналы могут иметь химическую природу. Известно, например, что если в аквариум с рыбками налить немного воды из другого аквариума, в котором стайка таких же рыбок недавно спасалась от хищника, то подобная паника может охватить и обитателей первого аквариума, В данном случае информация передается с помощью особого вещества — феромона*) тревоги.
В анонимном сообществе первыми обычно издают сигналы особи, оказавшиеся на периферии. Поскольку члены стаи все время меняют свое положение, то невольным лидером, за которым они устремятся, может оказаться любое животное, Такое сообщество называют эквипотенциальным.
Анонимные сообщества, члены которых не проявляют агрессии по отношению к вновь присоединившимся особям своего вида, получили название открытых. К ним относятся, например, стада жирафов или кенгуру, которых привлекает вид сородичей, но они с равной легкостью присоединяются к данной группе, а затем покидают ее.
Другой тип анонимного сообщества — закрытое. Его члены не различают друг друга "персонально", но могут выделять животных, принадлежащих к другому сообществу ("чужаков"), например по запаху. К типичным закрытым анонимным сообществам относятся колонии крыс-пасюков (Rattus norvegicus). При появлении на территории колонии чужака все ее взрослые члены набрасываются на него и либо убивают, либо изгоняют. Единственный признак, по которому крысы отличают "своих" от "чужих", — это специфический для каждой колонии запах. Если крысу потереть подстилкой, взятой из другой колонии, она сейчас же будет убита сородичами, с которыми до этого жила в полном мире (рис. 5.2).

Рис. 5.2. Агрессивное столкновение между крысами-пасюками
Более высокий тип общественной организации — это индивидуализированные (персонифицированные) сообщества, т.е. объединения животных с упорядоченной структурой, в которых каждый член "знает" всех остальных "персонально" ("в лицо").
Стабильные замкнутые группировки, основанные на устойчивых отношениях между составляющими их особями, характерны для представителей отряда хищных млекопитающих (см.: 5.5.2), особенно тех, кто добывает пищу коллективной охотой (волки, гиеновые собаки и др.), а также для многих видов птиц и приматов. Ядром такой стаи обычно бывает группа достаточно опытных, немолодых животных, которые давно знают друг друга и находятся в "дружеских" отношениях. Для подобных групп характерно сочетание проявлений типичных черт иерархии доминирования (см.: 5.3.1) с более сложными "личными" отношениями между отдельными особями.
Другой важной особенностью таких групп является забота старших особей о целостности сообщества и безопасности его членов. Таким образом, при анализе структуры сообществ подобного типа можно выявить и черты иерархии доминирования с неизбежными проявлениями агрессии, и проявление феномена "ролей", когда лидерство попадает к наиболее опытным членам группы, распределяющим эту обязанность между собой (подробнее о "ролях" см.: 5.3.4 и 5.3.7). В целом для таких сообществ характерна очень высокая пластичность индивидуальных контактов, включая черты неформальной привязанности и "дружбы". У высших позвоночных (приматов, дельфинов) сообщества имеют некоторые черты открытых групп (отсутствие агрессии по отношению к чужакам), которые сочетаются с очень сложными и устойчивыми взаимодействиями между особями. Примеры поведения высокоорганизованных животных в сообществах "индивидуализированного" типа рассмотрены в разделах 5.5.2—5.5.3.
Существуют разные подходы к оценке уровня сложности социальных контактов и относительной роли агрессии и "дружелюбия" в индивидуализированных сообществах. Многочисленные исследования были проведены в зоопарках и в экспериментальных лабораторных вольерах. При этом неизменно возникал вопрос о том, насколько отношения между особями в таких группах действительно соответствуют тому, что происходит в природе, в естественных группировках животных. Критический анализ этой проблемы можно найти в книгах О. Меннинга (1982; Manning, Dawkins, 1992).
Более прямой ответ на этот вопрос дают многочисленные наблюдения за природными популяциями в естественных для вида условиях. Исследователи постепенно приучали животных к своему присутствию, не вмешиваясь в жизнь группы. Так, Дж.Гудолл пишет (1992), что в своих взаимоотношениях с шимпанзе прошла три этапа. На первом этапе она старалась преодолеть естественный для диких животных страх, когда обезьяны при ее появлении разбегались. На втором этапе у обезьян возникали ориентировочно-исследовательские и агрессивные реакции, которые со временем угасли. И, наконец, наступило время, когда обезьяны стали встречать исследовательницу как сородича. При ее появлении они издавали специфический приветственный крик, раскачивали ветви деревьев, а в некоторых случаях вообще не обращали на нее внимания. Наблюдая за шимпанзе более 30 лет, Гудолл могла различать "в лицо" всех членов группы и даже иногда знала "биографии" отдельных особей от рождения и до смерти.
Исследования природных популяций человекообразных обезьян (Шаллер, 1968; Фосси, 1990; Гудолл, 1992) и некоторых других млекопитающих, в частности медведей (Пажетнов, 1985; 1991), гиеновых собак (Дж. и Г.Лавик-Гудолл, 1977) и др., а также птиц, например воронов (Хейнрих, 1994), позволили выявить, какие именно феномены, описанные при наблюдениях в неволе, действительно существуют и в естественных условиях, а какие черты социального поведения этих видов в неволе следует рассматривать как артефакт. Многие из этих работ опубликованы в виде не только научных статей, но и увлекательных научно-популярных книг, сочетающих научную достоверность с художественной формой изложения. Тем самым этологи 60—90-х годов продолжили традицию, начатую К. Лоренцем (1978; 1984; 1992; 1994) и Н. Тинбергеном (1970; 1974; 1978), чьи книги стали классикой.
Согласно общепринятым представлениям, в основе поведения животных в сообществе лежит взаимодействие двух факторов: 1) общественного инстинкта, побуждающего их объединяться с сородичами и поддерживать с ними постоянные контакты; 2) внутривидовой агрессивности, которая позволяет установить и поддерживать определенный порядок в сообществе. На значение третьего фактора — уровня развития рассудочной деятельности животных данного вида, который определяет сложность отношений в сообществах высокоорганизованных млекопитающих и птиц, — одним из первых указал Л.В.Крушинский (1973) (см.: 5.3.9). Впоследствии о значении этого фактора в установлении "неформальных контактов" между животными писали неоднократно.
Структурированность сообщества — поддержание иерархической организации, обеспечивается прежде всего благодаря феномену доминирования и подчинения.
Норвежский эколог Т. Шьелдерупп-Эббе (Schelderupp-Ebbe, 1924) обнаружил в группах домашних кур и уток строгую упорядоченность отношений между птицами. Каждая особь либо превосходит по силе партнера, либо уступает ему. Эти взаимоотношения были названы "порядком клевания". При формировании группы происходит "выяснение отношений" птиц друг с другом, в ходе которого постепенно выделяется одна, которая первой получает доступ к корму и гоняет от него всех остальных. Ниже ее на "лестнице доминирования" располагается птица второго ранга, которая превосходит всех, кроме главной, доминантной особи, и так далее. В самом основании находится особь, которую гоняют все члены группы.
Такая иерархическая система вырабатывается при столкновении птиц в борьбе за "ограниченный ресурс" (за место на насесте, пищу), и на ранних этапах ее установления происходит много драк. Однако когда иерархия устанавливается, она оказывается стабильной, поскольку порядок соподчинения особей устойчиво поддерживается. Обычно при приближении высокоранговой птицы подчиненные особи уступают ей без сопротивления.
Анализ показал, что предпосылкой стабильности иерархии является индивидуальное узнавание особей. В экспериментах, в которых одну и ту же птицу помещали последовательно в разные группы, "откалиброванные" таким образом, что она занимала в них разное иерархическое положение, куры обнаружили незаурядные способности к запоминанию и распознаванию членов каждой группы и без колебаний занимали причитающееся им положение.
У кур иногда устанавливается совершенная линейная иерархия, так что ни одна птица никогда не клюет особей, стоящих выше ее на иерархической лестнице (табл. 5.1). Подобные "идеальные" сообщества исключительно редки. Среди беспозвоночных их образуют, например, сверчки и речные раки, у которых иерархические отношения также строятся на основе индивидуального распознавания. В то же время у большинства видов животных обнаруживаются различные отклонения от строгого линейного порядка.
Описанное явление — формирование иерархической структуры группы — стало рассматриваться как механизм, благодаря которому одно или несколько животных получают приоритет во всех жизненных ситуациях.
Предполагалось, что иерархическое ранжирование выделяет наиболее жизнеспособных особей, обеспечивая преимущественный успех их потомства в процессе естественного отбора. Действительно, некоторые эксперименты прямо свидетельствовали о лучшей приспособленности членов жестко организованных сообществ. Например, в одних группах кур доминантную особь регулярно удаляли и заменяли незнакомой птицей, так что остальные вынуждены были постоянно заниматься "выяснением отношений", тогда как членов контрольных групп не тревожили. В контрольных группах агрессивные стычки происходили реже, яйценоскость кур была выше, т.е. группы с постоянным составом обладали явным преимуществом благодаря стабильной социальной среде. Аналогичную иерархическую структуру и свойства имеют колонии серых крыс (см.: 5.5.1).
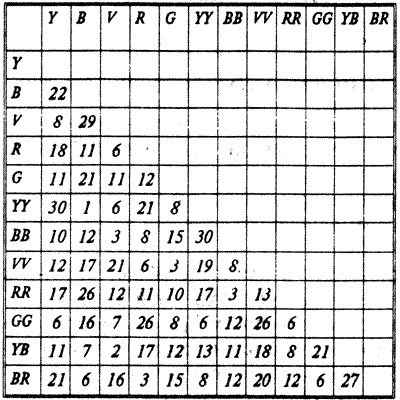
Таблица 5.1 "Идеальная" линия иерархии в группе, состоящей из 12 кур
Примечание. В опыте каждая из птиц была индивидуально помечена. Таблица составлена на основе этограмм регистрации контактов птиц между собой в течение определенного времени наблюдения. В вертикальных столбцах — число клеваний, которые данная курица нанесла другим членам группы, в горизонтальных — число клеваний, которые она же получила от других членов группы.
"Идеальность" данной иерархической структуры выражается в том, что ни одна курица не клюнула ни одну из особей, стоящих на иерархической лестнице выше ее.
Один из наиболее распространенных методических приемов, в особенности при исследовании физиологических основ доминирования и агрессивности, — это экспериментальный анализ поведения в "конкурентных ситуациях", когда моделируется ограниченность доступа к ресурсам. Для этого двум животным, на определенное время лишенным пищи или воды, одновременно открывают путь к одной кормушке или поилке. В зависимости от условий эксперимента подкрепление может получить либо только один из конкурентов, либо доминантом считается животное, которое больше съедает или дольше контролирует кормушку.
Эксперименты такого рода, проведенные на животных разных видов, показали, что корреляция между разными показателями доминирования (конкуренция за пищу, воду, территорию, доступ к половому партнеру, возможность уйти в гнездо и т.п.) может быть слабой или совсем отсутствовать. Даже в строго контролируемых лабораторных условиях при использовании генетически однородных однополых и одновозрастных животных не найдено "единого" доминанта.
Образование и поддержание стабильных общественных группировок невозможно без внутривидовой агрессии (см.: рис. 5.2). Она выполняет следующие функции:
1) обеспечение изоляции группы в пределах одной популяции, что, например, препятствует перемешиванию стай при встрече их на одной территории;
2) противодействие проникновению в данное сообщество чужих особей, а также ограничение числа производителей путем изгнания части самцов; в таких случаях возникает "резерв" мигрирующих особей, за счет которого пополняется недостаток производителей в других популяциях; у многих видов существование подобного резерва обеспечивает также обмен генами между группами;
3) участие в формировании и поддержании структуры индивидуализированных сообществ.

Рис. 5.3. Примеры истинных агрессивных конфликтов у млекопитающих (зебры)
Птицы, демонстрируя силу, распушают перья и расправляют крылья.
Иерархические отношения между животными устанавливаются в результате конфликтов — либо настоящих драк (рис. 5.3), либо угрожающих демонстраций, причем последние бывают не менее эффективными, чем первые (рис. 5.3, внизу). Как правило, они включают различные приемы, благодаря которым животные кажутся крупнее, чем они есть на самом деле. Например, хождение на двух ногах, вздыбливание шерсти, подскакивание шимпанзе действуют не менее эффективно, чем прямое нападение. Такие же элементы поведения обнаруживаются у ящериц (рис. 5.4). Птицы, демонстрируя силу, распушают перья и расправляют крылья.

Рис 5.4. Ящерицы рода Anolis для "устрашения" противника увеличивают свой объем, раздувая специальный мешок на шее (вверху), внизу показан силуэт того же животного в спокойном состоянии
К. Лоренц предположил, что существование групп с тесными индивидуальными связями между особями возможно только у животных с достаточно развитой способностью к направленной агрессии, у которых объединение двух или большего числа особей имело значение для выживания. Этой проблеме посвящена его книга "Агрессия (так называемое зло)", написанная в 60-е годы и переведенная на русский язык в 1994 г.
Логический анализ наблюдений за поведением животных при проявлениях агрессивности, страха или взаимного влечения позволил Лоренцу сделать заключение о существовании тонкого баланса между этими состояниями, а также о неотделимости проявлении агрессивности как центробежной тенденции, от проявлений "любви" — общественного инстинкта, которые обуславливают взаимное притяжение особей.
Написанная очень увлекательно, книга Лоренца не только внесла вклад в специальную область биологии — этологию, но и явилась одним из важнейших научно-философских произведений XX в. В ней приведены многочисленные доказательства того, что агрессивность является врожденным, наследственно обусловленным свойством всех высших животных. Прослеживая подобные свойства в поведении человека, К.Лоренц приходит к выводу, что "есть веские основания считать внутривидовую агрессию наиболее серьезной опасностью, какая грозит человечеству в современных условиях культурно-исторического и технического развития" (Лоренц, 1994). Это положение повлекло за собой обвинения автора в недооценке социальной природы человека. Оно способствовало отрицательному отношению к этой книге в СССР и на десятилетия задержало ее перевод на русский язык. Между тем, основное значение книги заключается прежде всего в изложении основ регуляции социальных отношений у животных.
Поддержание структуры сообщества осуществляется на основе агонистического поведения, т.е. комплекса реакций нападения, угрозы, подчинения и бегства. Как уже упоминалось, установление иерархии может происходить в результате прямых столкновений и драк, тогда как по мере ее упрочения в большинстве случаев прямые проявления агрессии уступают место угрозам и демонстрациям.
Выше (раздел 4.3) были рассмотрены типичные примеры угрожающих демонстраций у рыб и чаек. Согласно гипотезе Н.Тинбергена, выраженность практически всех демонстраций определяется соотношением уровня агрессивности и страха, которые в данный момент переживает животное (рис. 5.5 и 5.11).
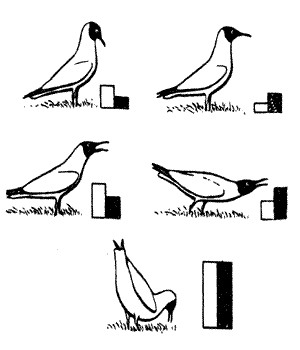
Рис.5.5. Позы угрозы у обыкновенной чайки, связанные с разным соотношением тенденций к атаке (темные столбцы) и к избеганию (светлые столбцы) (по Мак-Фарленду, 1988}
Знаменитая серия зарисовок немецкого этолога П.Лейхаузена показывает мимику домашней кошки при разных сочетаниях уровней агрессии и страха (рис. 5.6): агрессивная тенденция усиливается слева направо, страха — сверху вниз.
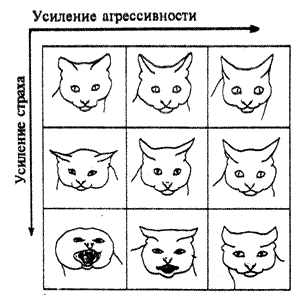
Рис. 5.6. Изменения мимики кошки в соответствии с разными уровнями тенденции к агрессии и реакции страха (по П. Лейхаузену)
Другую группу демонстраций составляют ритуализованные движения, тормозящие агрессию со стороны более сильных сородичей. Их называют позами "покорности" или "умиротворения". По всем своим параметрам они противоположны жестам угрозы. Так, коралловая рыбка, угрожающая сородичу, демонстрирует ему свой яркий наряд, распахивает плавники или жаберные крышки, движется резко, демонстрируя силу. Когда же она "просит пощады", все выглядит совершенно по-другому; ее окраска бледнеет, она прижимает плавники, поворачивается к сопернику узким сечением тела, стараясь казаться маленькой, движется медленно, крадучись. Подобным же образом, демонстрируя угрозу, птицы распушают перья, чтобы казаться как можно крупнее, тогда как в знак подчинения они прижимают перо, и их силует становится более тонким. У макаков-резусов позы и движется доминантного и подчиненного животных диаметрально различны (рис. 5.7 и 5.11).

Рис.5.7. "Стиль" поведения доминантных и подчиненных особей макаков-резусов (Меннинг, 1982)
Выразительные позы угрозы и подчинения у воронов (рис. 5.8) не оставляют сомнений в их функциональном "назначении".

Рис. 5.8. Ворон в позе угрозы и в позе подчинения (по Хейнриху, 1995)
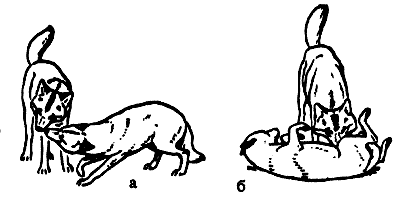
Рис.5.9. Позы подчинения у волков.
а — ритуальное выпрашивание пищи; б — переворачивание на спину (по Eibl-Eibesfeldt, 1977)
Формы ритуалов умиротворения многообразны, тем не менее некоторые из них (поза подставления, реакция выпрашивания корма) у самых разных групп животных одинаковы (рис. 5.9). К. Лоренц (1994) выделил несколько основных особенностей таких ритуалов:
1. Демонстративное подставление наиболее уязвимой части тела. Так, побежденный волк отворачивает голову и подставляет победителю чрезвычайно ранимую боковую сторону шеи, выгнутую навстречу укусу. Галка подставляет под клюв птицы, которую нужно умиротворить, свой незащищенный затылок — обычную мишень при серьезном нападении с целью убийства. У многих видов птиц оперение на таких участках тела имеет особую окраску. У галок оно шелковисто-серое, а у тех врановых, которые крупнее и темнее на затылке имеется более светлое пятно. По данным Н.Тинбергена (1974), чайки, демонстрируя умиротворение, отворачивают от противника клюв (см.: рис. 4.6 в), при этом привлекает внимание их белоснежный затылок (как у обыкновенной чайки) или особый рисунок на светлом фоне (как у молодой моевки).
2. Воспроизведение некоторых элементов детского поведения. Распространенный вариант ритуала умиротворения у птиц — это имитация позы птенца, выпрашивающего корм (рис. 5.10).
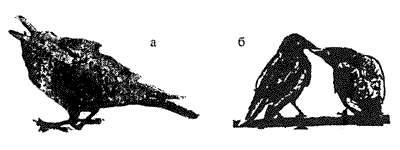
Рис 5.10. Ритуал "выпрашивания корма" у воронов а — рисунок (по Хейнриху, 1995), б — фотография Хейнриха
В репертуаре социального поведения псовых жесты умиротворения также играют важную роль; они происходят непосредственно из отношений щенка с его матерью и являются производными от движений сосания или просьбы накормить (см.: рис. 5.9). Как предполагает К. Лоренц (1994), эти действия превратились в ритуалы у тех животных, у которых изначально существовали более общий, социальный смысл, оказалось возможным формирование крупных социальных групп на базе супружеских пар и семей.

Рис.5.11. Сигнальные позы у клуши
а — "прямая" поза, означающая угрозу; б — "сгорбленная" поза умиротворения у той же птицы, при которой голова опущена, клюв направлен вверх, а крылья плотно прижаты к туловищу, так что запястные суставы, демонстрирующиеся при угрожающей позе, оказываются незаметными (Меннинг, 1982}
3. Выражение социальной покорности с помощью действий, характерных для поведения самки при спаривании. Эти действия типичны для многих обезьян, особенно павианов. Ритуальный поворот задней части тела означает признание более высокого ранга другой обезьяны. Позу подставления они выполняют с самого раннего возраста без всякого обучения, даже при воспитании в изоляции от сородичей (рис. 5.12). Как уже упоминалось, те части тела, которые животные демонстрируют в знак угрозы или подчинения (релизеры — см.: 3.4), окрашены особенно ярко и заметно, чем подчеркивается выразительность церемонии. Так, у некоторых видов павианов задняя часть тела, которую они демонстрируют противнику в позе подставления, раскрашена самым фантастическим образом.
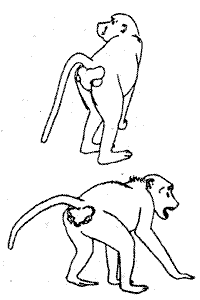
Рис 5.12 Поза "подставления у самца павиана занимающего "подчиненное" положение
Представление о том, что в условиях ограниченного доступа к какому-либо ресурсу доминирование способствует его получению в первую очередь, подтверждено в многочисленных исследованиях социального поведения животных (подробнее см.: Резникова, 1997).
С ростом числа исследований на животных разных видов выявилось множество отклонений от классической линейной схемы иерархической структуры сообществ. Наиболее распространенное отклонение — превращение "лестницы" или "цепочки" в двухуровневую структуру: доминант — и все остальные. Случаи такого абсолютного доминирования наблюдаются, например, при содержании самцов домовых мышей или самцов колюшки в ограниченном пространстве. В иных случаях в иерархических "цепочках" могут наблюдаться "петли" или "треугольники", когда особь А гоняет особь В, особь В — особь С, но А боится С. Описаны даже такие ситуации, когда длинная иерархическая цепочка замыкалась в круг.
Показано, что в сообществах могут действовать параллельные системы иерархии — у самцов и у самок. Изменение ранга самки, как это было описано К. Лоренцем (1978) при изучении сообщества галок, может происходить, если она становится супругой высокорангового самца. У большинства видов самцы имеют более высокие ранги, однако имеются и исключения.
У некоторых видов животных внутри большого сообщества существуют стабильные группировки — кланы, чаще всего образованные одной или несколькими самками с их потомством одного-двух поколений (см. Крук, 1977) Такие "материнские кланы" можно наблюдать, например, у слонов (И. и О.Дуглас-Гамильтон, 1981), дельфинов (Прайер, 1981, см. также обзор. Крушинская, Лисицына, 1983), гиеновых собак (Дж. и Г. Лавик-Гудолл, 1977). На рис. 5.13 приведена схема "построения" группы павианов при перемещении на открытом пространстве.
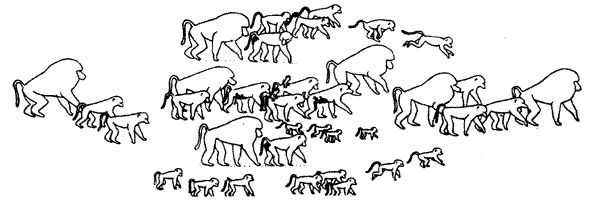
Рис 5.13 Стая павианов на марше через открытую равнину
Самые крупные животные с большими гривами — половозрелые самцы
Самки с детенышами держатся в наиболее безопасной зоне — в центре группы
на схеме изображены и самки в эструсе (у них зачернена задняя часть тела), они находятся рядом с доминантными самцами (по De Vore, 1965)
Помимо относительно стабильных группировок и кланов, члены сообщества могут образовывать и временные коалиции, направленные либо на достижение "сиюминутной выгоды", либо на то, чтобы повысить свой общий ранг. Множество примеров подобных ситуаций дают исследования приматов (японских макаков, зеленых мартышек, павианов, шимпанзе). Так, два—три самца могут объединиться, чтобы отвлечь доминанта, инсценируя драку или интересную находку, и, пока доминант "наводит порядок", по очереди получить доступ к пище или даже увести самку. Пример такой же "эксплуатации" молодым самцом принятого в сообществе доброжелательного отношения ко всем детенышам, можно видеть на рис. 5.14.
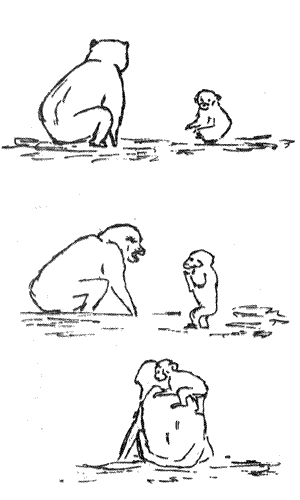
Рис. 5.14. Неполовозрелый самец варварийской обезьяны (берберийского макака) и детеныш этого вида. Сначала они сидят недалеко друг от друга (вверху), Затем самец подманивает детеныша, делает ему дружественное гримасу, детеныш к нему подходит (в середине), забирается к самцу на спину, и тот может отнести и "предъявить" его старшему самцу. Благодаря этому он может остаться рядом с доминантом в относительной безопасности
Существует достаточно много примеров, когда в сообществе приматов складывается не столько иерархия особей, сколько иерархия группировок (Гудолл, 1992).
Особенности структуры сообществ, выявленные в наблюдениях за животными разных видов, показали, что внутривидовая иерархия, ее структура и механизмы поддержания, а также возможные отклонения зависят от ряда факторов.
Конкретная ситуация в сообществе почти всегда лабильна и также может выходить за рамки простых иерархических отношений. Этологи выделяют некоторые правила, соблюдающиеся в группах животных в обход существующей иерархической системы. Одно из таких правил — так называемое "право обладания". Так, например, если низкоранговая особь шимпанзе добывает редкое лакомство, все остальные окружают ее, выпрашивая свою долю. Высокоранговые животные при этом не пользуются никакими преимуществами.
Пластичность поведения животных в сообществе обусловлена их богатым индивидуальным и социальным опытом. Создается впечатление, что классическая линейная иерархия является лишь некой врожденной "матрицей", характерной для многих общественных животных. В течение жизни на нее накладывается приобретенное умение оценивать и использовать социальные связи.
Это положение было подтверждено экспериментально (Anderson, Mason, 1978). Были сформированы две группы макаков-резусов: одна состояла из обезьян, выращенных в изоляции, с "искусственными" матерями (как в известных опытах Harlow et al, 1963), а члены другой росли со своими матерями и сверстниками. В обеих группах экспериментаторы искусственно создавали конкуренцию за бутылочку с водой. Оказалось, чтo у животных, выращенных в изоляции, устанавливается стабильный порядок доступа к бутылочке, напоминающий "порядок клевания" у кур. В то же время у животных с нормальным опытом социальных контактов была обнаружена достаточно высокая лабильность социального поведения. Они были, например, способны объединяться в коалиции, могли искать защиты у доминанта в спровоцированной ими же драке и тем самым отвлечь его от бутылочки и т.п. Подобные закономерности обнаружены и у беспозвоночных, например в опытах В.А.Харькива (1993) на муравьях (подробнее см.: Резникова, 1997).
Форма общественной организации может варьировать у данного вида в зависимости от местообитания и плотности популяции. Существует ряд четких примеров изменения структуры сообщества в зависимости от плотности населения. Часто перенаселение ведет к появлению в сообществе более выраженной иерархии. В экспериментах, проведенных на таких разных животных, как луна-рыба и мышь, было обнаружено, что в условиях высокой плотности самцы образуют типичную иерархию, но когда позволяет пространство и существуют укрытия, они переходят к территориальности.
Зависимость формы организации сообществ, в частности размера групп, от экологических условий лучше всего исследована у млекопитающих. Однако суммировать весь ряд факторов, влияющих на величину группы и характер внутригрупповых взаимоотношений, далеко не просто. На первый план выступают два фактору: распределение пищевых ресурсов и давление со стороны хищников.
Если ресурсов питания много, то это способствует образованию больших групп. Например, у пасущихся на равнинах травоядных большие группы мало угрожают сохранению пищевых ресурсов. Напротив, виды, находящиеся на вершине пищевой цепи (такие как волк или крупные кошки), не могут существовать при высокой плотности.
Что касается второго фактора — угрозы со стороны хищников, то очевидно, что большим группам справляться с этой угрозой легче.
Демографические параметры популяции, влияющие на характер взаимоотношений особей в группах, зависят от обоих этих факторов. Это относится в первую очередь к числу взрослых самцов в группах: в областях с большим давлением со стороны хищников оно заметно выше (подробнее см.: Резникова, 1997).
Одно из проявлений пластичности структуры сообществ животных — ее изменения в зависимости от степени урбанизации среды. Воздействие антропогенных факторов может существенно влиять на пищевые ресурсы и наличие мест для гнездования птиц, меняя соответствующим образом размер и способ организации сообществ. Ярким примером таких влияний могут быть изменения, которые претерпели сообщества многих видов врановых птиц. Например, галки (Corvus monedula), типично колониальный вид, могут селиться разреженно и даже образовывать колонии ленточного типа. Напротив, представители одиночно гнездящегося вида — серые вороны (Corvus cornix) — в городах при изобилии корма и гнездового материала строят гнезда в непосредственной близости друг от друга, а в единичных случаях — даже на одном дереве (Константинов, 1992).
Говоря о пластичности социального поведения птиц, нельзя особо не упомянуть о вОроне (Corvus corax, L.). Ворон считался чисто одиночным видом, однако сейчас показано, что вороны могут сотнями кормиться на свалках и других массовых источниках корма (Константинов, 1992). Природа таких стай может быть очень сложной, и их нельзя считать простыми скоплениями в указанном выше смысле (см.: 5.2.2). Американский зоолог Б. Хейнрих (1994) в течение многих лет наблюдал за поведением воронов в лесах штата Мэн. Приманками для них служили коровьи туши или специально разложенные куски мяса. Стаи воронов (он назвал их "ордами"), насчитывавшие от 15 до 300 птиц, постепенно расклевывали эти приманки. Наблюдения позволили установить, что такая концентрация воронов у приманок — явление совершенно особое; эти объединения отличны по своей природе от стай других врановых, которые кормились тут же. В частности, голубые сойки (Суапосttа cris-tata, L.) применяют альтернативную стратегию и, обнаружив корм, стараются скрыть это, чтобы не делиться с сородичами.
Самая главная особенность "орд" воронов состоит в том, что это не пассивные скопления. В поисках корма они летают парами или поодиночке и, обнаружив его, специальными криками ("воплями") широко извещают об этом, как бы приглашая всех желающих присоединиться. Эти крики нельзя услышать практически ни в какой другой ситуации. Хейнрих предполагает, что они развились из звуков, которые издают птенцы, выпрашивая корм.
Детальный анализ, который мы не будем приводить подробно, показал, что "приглашающие вопли" издают только молодые птицы. Благодаря этому они получают или удерживают доступ к корму, до которого их иначе не допустили бы владельцы территории - взрослая пара воронов. Хейнрих предполагает, что, помимо обеспечения пищей, у этих сборищ молодых птиц есть и другое предназначение — обретение социального статуса и "помолвки". Ворон, дока не имеющий пары, найдя корм, приглашает других птиц присоединиться к нему и тем самым не только получает или сохраняет доступ к пище, но, возможно, повышает свой статус, демонстрируя способности будущего кормильца при выведении потомства.
Этологические и генетические исследования показали, что в сообществах мышей иерархическая структура лабораторной популяции определяет преимущественное отцовство доминанта (McClearn, DeFries, 1972).
Однако, по данным исследований индивидуализированных сообществ высокоразвитых животных, связь между иерархическим рангом и числом оставленных потомков бывает далеко не очевидной. Подчиненная особь может успешно адаптироваться к поведению доминанта и получать "все блага жизни", даже не вступая в борьбу. У самца благородного оленя описана удивительная "клепто-репродуктивная стратегия". Молодой самец успешно спаривался с самками, пока хозяин "гарема" отвечал на вызовы других самцов.
Усложнение схемы иерархического строения сообщества связано и с "распределением ролей" (или "разделением труда") животных в группе. Оно описано у некоторых совместно охотящихся хищных рыб (тунца, макрели), а также у ряда видов млекопитающих, таких как бобры, львы, волки, гиены, гиеновые собаки, шакалы и др. Этот феномен представляет собой выполнение членами группы различных, но четко определенных по функции действий, например при охоте или охране территории. Возможность выполнения разных ролей в сообществе определяется у каждой особи сложнейшим сочетанием видовых, наследственных ФКД и поведения, основанного на индивидуальном и социальном опыте. На "результирующей" влияния опыта и врожденных задатков и основана роль каждого индивидуума в сообществе.
В целом, "разделение труда" в сообществах животных, несомненно, делает их социальные отношения более сложными и многообразными. В разных ситуациях на первый план могут выходить особи, более способные к тому или иному виду деятельности. Такие отношения называют "ролевой иерархией".
Замечательное разделение труда существует в сообществе бобров. Обитающая в хатке группа животных выделяет "дежурных", которые по очереди следят за бобрятами. Они постоянно играют роль "спасателей", так как детеныши еще не очень хорошо плавают и, покидая хатку, могут не найти входа в нее, задохнуться в воде либо погибнуть на берегу. Бобры, работающие вне хатки, также выполняют разные функции, такие как снабжение бобрят пищей, охрана или строительство.
Существуют наблюдения, свидетельствующие о том, что у некоторых видов "разделение труда" включает в себя манипуляции поведением одних особей со стороны других. Наиболее известны, хотя и не полностью убедительны, знаменитые эксперименты К. Моуэра (Mower, 1940): в камеру помещались несколько крыс, которые могли нажимать на рычаг для получения пищи, однако кормушка находилась в отдалении от него. Оказалось, что в такой ситуации на рычаг нажимали лишь немногие особи, обеспечивая пищей всех остальных. Остается неясным, почему "работала" только часть животных, а другие вели "паразитический" образ жизни.
Другой, более убедительный пример "разделения труда" и "эксплуатации" в группе молодых шимпанзе описал Л.А. Фирсов (1977). Он провел свои наблюдения на озерном острове в Псковской обл., где молодые животные летом вели свободный образ жизни. На первом этапе эксперимента шимпанзе могли получить приманку из открывающегося ящика только в результате успешных действий партнера. Когда они усвоили этот навык, отношения между ними начали изменяться. Достаточно сбалансированные в начале эксперимента, они быстро переходили в фазу "беззастенчивой эксплуатации", когда большую часть заработанной приманки получали не "работники", а "наблюдатели". Прежняя картина отношений восстанавливалась лишь после энергичных столкновений. Несомненно, такое "разделение труда", особенно путем манипуляций поведением партнеров, требует известного социального опыта и гибкости поведения. Сходные отношения складывались и в группе молодых ворон (в начале эксперимента их возраст составлял 2 месяца), которые обучались нажимать на рычаг для получения корма одновременно. И в этом случае доля пищедобывательных реакций, совершенных членами группы, с самого начала была различной, и это различие углублялось в процессе эксперимента. В конце концов около 80% всех реакций совершала одна из птиц, получая при этом лишь 38% заработанных ею порций корма. Остальные 3 птицы получали в среднем по 20% всех подкреплений, даже если совершали не более 2% нажатий на рычаг. Существенно отметить, что взаимодействия между птицами были почти лишены признаков агрессивности, во всяком случае, ворона, нажимавшая на рычаг, не пыталась отгонять других членов группы от "заработанного" ею корма. Возможно, это происходило потому, что самыми активными "добытчиками" были "низкоранговые" особи (Зорина, 1996). Таким образом, в данной ситуации представители весьма разных групп позвоночных ведут себя довольно сходным образом. Изучение индивидуализированных сообществ животных дает много примеров крайне важной роли сотрудничества для успешной деятельности и выживания группы. Наиболее яркие из них — это совместная охота у многих крупных хищников, совместное сложное строительство и выращивание потомства у далеких друг от друга видов, например у бобров, голых землекопов (см.: 5.5.1) и общественных насекомых — термитов, ос, пчел, муравьев (см.: 5.6.5).
В приведенных примерах сотрудничество между членами сообщества составляет видоспецифическую черту их поведения, непременно характерную для всех особей. Наряду с этим сотрудничество может возникать как индивидуальное приспособление отдельных особей к конкретным условиям среды, которое не предусмотрено видовыми стереотипами поведения. Такой вид сотрудничества представляет особый интерес для характеристики разумных компонентов в поведении животных (см.: Крушинский, 1986).
Наиболее убедительные примеры этих высших форм кооперации обнаружены у человекообразных обезьян. Так, Дж. Гудолл (1992) приводит в своей книге ряд примеров, когда шимпанзе оказывали помощь другим членам группы, которые могли и не быть их родственниками. Еще один пример — наблюдение Р. Футса за "говорящей" обезьяной Уошо (см. также: 5.4.2.2), которая жила на островке, окруженном рвом с водой и изгородью с электрическим током. Однажды молодая самка Синди, решив навестить Уошо, каким-то образом преодолела эту изгородь, но упала в воду и начала тонуть. Увидев это, Уошо, держась за траву, вошла в воду и умудрилась схватить Синди за руку, когда та, может быть, в последний раз всплыла на поверхность. Следует подчеркнуть, что между обезьянами не было родственных отношений, да и познакомились они совсем незадолго до события (рис. 5.15).

Рис. 5-15. Спасение утопающего (рисунок В. Маркина)
Разбирая этот и подобные довольно многочисленные примеры, Дж. Гудолл полагает, что отбор родичей может играть роль в формировании способности к взаимопомощи не только среди родственных, но также и посторонних друг другу особей.
О способности к сотрудничеству не только у приматов, но и других высокоорганизованных позвоночных свидетельствуют упомянутые выше данные о врановых птицах. Своеобразным экспериментальным доказательством ее существования могут служить также данные опытов Л.С. Бондарчука, описанные в работе Крушинского и др. (1982), по одновременному обучению двух ворон в специальной камере, разделенной пополам прозрачной перегородкой. В каждой половине находилась педаль, при нажатии на которую появлялся корм, но это происходило только тогда, когда вороны нажимали на "свои" педали одновременно. Независимо от этого ворон научили открывать дверцы примыкавших к обоим отделениям камеры дополнительных клеток, где птицы видели кормушку с мясом. Каждый из навыков вырабатывался достаточно легко. Затем в главном эксперименте одну из ворон запирали в клетку, так что вторая птица при нажатии на педаль корма не получала.
Оставшись в одиночестве, ворона в конце концов открывала клетку и выпускала партнера. Этот факт рассматривается как доказательство способности птиц к экстренной интеграции независимых навыков в новой ситуации, т.е. к одному из относительно простых видов рассудочной деятельности (Зорина, 1997).
Анализируя проблему взаимопомощи и сотрудничества у высших позвоночных, Л.В.Крушинский (1986) приходит к выводу, что основным механизмом, обеспечивающим возможность таких взаимоотношений между членами сообщества, может быть только достаточно развитая рассудочная деятельность. Действительно, поведение Уошо в приведенном примере включает экстренную оценку новой, ранее никогда не имевшей места ситуации, способность поставить себя на место обезьяны, терпящей бедствие, и принять адекватное данной ситуации решение.
Развитие социального поведения животных в онтогенезе зависит, по крайней мере, от двух основных факторов. Первый из них — это генетически детерминированная видоспецифическая "система общения", т.е. система коммуникации с особями своего вида, основанная на обмене сигнальными позами, жестами, движениями, включая демонстрацию особых сигнальных структур релизеров (см.: 3.4). Как само продуцирование подобных сигналов, составляющих язык животных, так и их опознание, как правило, не требуют индивидуального опыта: оно относится к категории врожденных реакций. Второй фактор — существование определенных периодов индивидуального развития, в течение которых с наибольшей легкостью и быстротой формируются связи между особями: запечатление детьми внешнего облика родителей, которое будет определять общественные взаимодействия этих особей по достижении ими взрослого состояния (см. также: 3.5 и 7.1).
Важнейшая роль в становлении социального поведения (как и практически любых других его форм) принадлежит играм (см. обзор: Зорина, 1998). Особенно ярко роль игры проявляется у животных, которые живут в сложно организованных сообществах. В ходе игры используются элементы агонистического поведения и закладываются основы иерархических отношений между их участниками. По мере взросления игры многих животных приобретают все более грубый характер и нередко заканчиваются агрессивными эпизодами. Благодаря этому животные не только получают сведения о сильных и слабых сторонах своих партнеров по игре и об относительном иерархическом положении своей матери и матерей товарищей по играм, но и учатся драться, угрожать, устанавливать союзнические отношения. Это позволяет им впоследствии успешно конкурировать с другими членами сообщества, в котором способность отстоять свои права и повысить ранг часто зависит от умения драться.
Установлено, что в характере социальных игр четко отражается видовая специфика поведения взрослых животных. Как отмечает К.Э. Фабри (1993), у одних видов грызунов (морские свинки) игровая борьба отсутствует, а социальные игры сводятся к сигналам "приглашения". В отличие от них, у большинства других грызунов распространены именно игровые схватки. Особенно отчетливо игровые взаимодействия грызунов проявляются в физически сложной среде обитания (Мешкова, Федорович, 1996). Социальные игры очень характерны для хищных млекопитающих. В качестве примера современных исследований этого аспекта проблемы можно привести данные многолетних наблюдений Н.Г. Овсянникова (1993) за поведением и социальной организацией песцов (Alopex galopus, L.). Они свидетельствуют, что взаимодействия молодых песцов в процессе игры действительно обеспечивают механизмы социальной интеграции в выводках этих животных. Игровая борьба у песцов феноменологически не имеет ничего общего с настоящей агрессией, хотя отдельные движения могут быть похожими. В целом схватки зверьков в процессе игры производят впечатление более стереотипных, монотонных действий, чем при настоящих схватках. Автор приводит ряд доказательств того, что игровая борьба эмоционально положительна и оказывает на выводки интегрирующее влияние. По свидетельству Овсянникова, во время игры стираются различия социального положения и роли в сообществе, временно ослабевает психосоциальный стресс, который неизбежен при взаимодействиях по необходимости — для выращивания потомства, добывания пищи и т.п.
Соотношение игровой борьбы, подвижных и охотничьих игр у разных видов также различно. При этом, как отмечает К.Э. Фабри (1993), нужно учитывать, что сами по себе элементы социального поведения представляют собой ФКД, которые проявляются в "готовом" виде и не требуют тренировки для своего созревания. Роль игры в становлении социального поведения состоит в том, что, если на ранних этапах она складывается из отдельных компонентов, то по мере взросления эти компоненты все больше интегрируются в единое целое.
Один из вариантов социальных игр — игры матери с детенышем. Они характерны для хищных млекопитающих, но особенно развиты и выражены у человекообразных обезьян: мать играет с детенышем с первых же месяцев жизни и до окончания подросткового периода.
В процессе онтогенеза у многих животных меняются образ жизни и соотношение разных типов контактов. Так, например, молодые лисицы и медведи живут семейными группами, в которых проявляются многообразие и сложность дружелюбных взаимодействий, а став взрослыми, ведут одиночный образ жизни. При этом у взрослых бурых медведей дружелюбные контакты, свойственные молодым, полностью уступают место выраженной агрессивности по отношению друг другу (Пажетнов, 1985; 1991).
Наиболее сложный процесс развития проходит социальное поведение человекообразных обезьян. На рис. 5.16 показано, как могут играть молодые гориллы.
По наблюдениям Гудолл (1992), детеныши шимпанзе имеют достаточно долгий подростковый период, когда в тесных контактах с сородичами они овладевают не только многочисленными пищедобывательными и оборонительными навыками, но усваивают и основные паттерны коммуникативных реакций, и правила поведения в сообществе.
5.3.9. ВЛИЯНИЕ УРОВНЯ РАЗВИТИЯ ЭЛЕМЕНТАРНОЙ РАССУДОЧНОЙ ДЕЯТЕЛЬНОСТИ НА СПЕЦИФИКУ ОБЩЕСТВЕННЫХ ОТНОШЕНИЙ ЖИВОТНЫХ
Л.В. Крушинский (1973) одним из первых систематизировал данные о том, что степень многообразия и пластичности отношений в сообществе животных тесно связана с уровнем их психического развития. В своей монографии "Биологические основы рассудочной деятельности" он рассмотрел специфику общественных отношений у животных, обладающих высокоразвитой рассудочной деятельностью, — врановых, дельфинов, хищных млекопитающих (псовых, кошачьих, медведей) и разных видов приматов, сопоставив ее с уровнем развития их рассудочной деятельности по экспериментальным данным и по наблюдениям в природе (Крушинский, 1986. С. 176-208).
Ниже (см.: 5.4) рассмотрена организация сообществ этих животных в других аспектах, не дублирующих содержание упомянутого раздела книги Крушинского, к которой мы и отсылаем читателя. Л.В. Крушинский пришел к выводу, что в сообществах высокоорганизованных животных взаимопомощь и сотрудничество играют весьма существенную роль, даже более значительную, чем агрессивность, связанная с поддержанием иерархической структуры. Речь идет не о том, что этот фактор определяет тип организации сообщества: независимо от уровня развития рассудочной деятельности виды могут вести как групповой, так и одиночный образ жизни. На основе приведенных данных Л.В. Крушинский (1986) делает вывод о том, что у наиболее умных животных из рассмотренных им групп — обезьян и дельфинов — наблюдаются и наиболее сложные формы социальных взаимоотношений (см. также: 5.5.3). Они не укладываются в рамки простых территориально-иерархических отношений, формирующихся на основе агрессии. У хищных млекопитающих, по его оценкам характеризующихся более низким уровнем рассудочной деятельности, социальные связи также относительно менее сложны (см.: 5.5.2).
Рис. 5-16. Разные типы игр молодых горилл (по Шаллеру, 1963)
Наиболее полные и убедительные конкретные доводы в пользу такого представления приводит Дж. Гудолл (1992. С. 427—428). Обобщая огромный объем собственных наблюдений за социальными взаимодействиями шимпанзе, она пишет, что это именно та сфера приспособительной деятельности животного, в которой требуются хорошее понимание причинно-следственных связей, мобилизация всех самых сложных познавательных способностей для достижения успеха и поддержания своего социального положения. Так, при рассмотрении вопроса о возрастных изменениях иерархического статуса самцов она указывает, что в ряде случаев борьба за доминирование напоминает "состязание характеров, в котором большое значение имеют ... изобретательность и упорство". В ее работе приводятся многочисленные примеры того, как низкоранговая особь достигает желаемой цели с помощью хитроумных обходных маневров, даже при явном неодобрении старшего по рангу. Это требует умения планировать свои действия и манипулировать поведением сородичей (см. также: 5.5.3), что как раз и относится к сфере разумного поведения.
Классическим примером роли такой сообразительности в достижении доминантного положения стал самец Майк, который догадался использовать пустые металлические канистры для усиления эффекта своего угрожающего поведения (Гудолл, 1992). Шимпанзе учитывают тот факт, что положение в группе может улучшиться, если "заключить дружеский союз" с другим самцом.
Соотношение сил в группе шимпанзе постоянно меняется, и каждый из них должен всегда быть настороже, уметь оперативно учитывать особенности сиюминутной ситуации и мгновенно менять в соответствии с ними свое поведение, иначе может последовать суровое возмездие. Как пишет Гудолл, молодой самец, уже направившийся к самке и начавший демонстрировать свои намерения, немедленно остановится и примет нейтральную позу, если на сцене появится самец более высокого ранга. Наблюдения показывают, что шимпанзе оценивают структуру сообщества отнюдь не только по результатам прямых агрессивных взаимодействий. Благодаря наблюдениям за контактами отдельных особей шимпанзе "вычисляет" на этой основе полную картину и собственное положение в ней: "если А гоняет В, а В угрожает С, следовательно, С ниже рангом, чем А". Практика показывает, что шимпанзе способны к такого рода умозаключениям. Эту сторону поведения некоторые авторы называют "social cognition". Именно подобные факты послужили основанием для высказанного американским исследователем Примэком (Premack, 1983) предположения о том, что у шимпанзе есть такая форма дедуктивного мышления как способность к транзитивному заключению, и побудили изучать ее в физиологическом эксперименте.
Не менее важную роль может играть и присущее шимпанзе умение понимать и оценивать выразительные движения, а также знания, намерения и другие внутренние (умственные) состояния сородичей, строя в соответствии с этим свое поведение. Тесно связанное со способностью к самоосознанию умение мысленно поставить себя на место другого и предугадать его действия Примэк (Premack, 1978; 1983) рассматривал как одно из наиболее сложных проявлений познавательной деятельности животных. В ряде работ показано, что такой способностью действительно обладают только человекообразные обезьяны. Так, Сифард и Чини (1993), исследуя язык восточно-африканских верветок (см. также: 5.4.3.3), задавались вопросом, могут ли обезьяны понимать разницу между своими собственными представлениями и знаниями и представлениями других особей, а также могут ли они специально проинформировать о чем-то сородича, или изменить неверные представления у другого.
Чтобы ответить на эти вопросы, авторы провели эксперименты с группами макак резусов и японских макак, обеспечивая некоторых особей информацией, которой другие не владели. Особенно красноречивыми оказались результаты экспериментов, в которых мать имела возможность "сообщить" своему детенышу о местоположении корма и о появлении хищника. По условиям опыта мать прекрасно видела, что детеныш не осведомлен, например, о грозящей ему опасности. Оказалось, однако, что у низших обезьян мать никак не пытается воздействовать на поведение детеныша, и, по-видимому, эти животные не принимают в расчет намерения сородичей.
Это вполне соответствует поведению низших обезьян в природе. Например, детеныши верветок, начиная издавать крики тревоги или реагируя на сигналы других, делают много ошибок. Некоторые из них безобидны: например, детеныш издает крик "орел", видя пролетающего над головой голубя. Другие ошибки могут стоить детенышу жизни, например, когда он смотрит вверх, услышав сигнал о приближении змеи. Однако исследователи не обнаружили доказательств того, что взрослые "поправляют" детенышей или как-либо поощряют поведение тех, кто издает сигналы правильно и адекватно на них реагирует. Детеныши верветок учатся только посредством наблюдения, по-видимому, в связи с неспособностью взрослых особей оценить, что уровень знания детенышей уступает их собственному. Эти результаты перекликаются с данными, полученными Д. Повинелли с соавт. (Povinelly et al., 1992) в остроумном эксперименте с "переменой ролей". Обезьянам (макакам и шимпанзе) предлагали игровой аппарат, напоминающий детский настольный хоккей, в котором партнеры играют попарно и по очереди дергают за рычаги. В данном эксперименте один из двух партнеров (по одну сторону аппарата — животное, по другую — партнер, человек), "информатор", видел, где спрятана приманка, но достать ее не мог (его рычаг был заблокирован), а мог лишь указать на нее второму партнеру — "оператору", который, наоборот, не видел, где находится приманка, и мог ее достать, лишь пользуясь указаниями "информатора" и дергая соответствующий рычаг. Оказалось возможным научить макак хорошо выполнять свои роли: одни животные становились "информаторами", другие — операторами. К перемене ролей они оказались совершенно не способными. Зато шимпанзе прекрасно поняли, что на месте партнера нужно выполнять не свою прежнюю роль, а ту, что на этом месте выполнял человек. Это означает, что шимпанзе, выполняя роль информатора, "влезали в шкуру" оператора, примеривались к его роли и сознавали, что партнер может, а чего не может делать, и наоборот. Поэтому шимпанзе, в отличие от низших обезьян, легко справлялись с задачей перемены ролей. Сходные данные о различиях в степени осознания своих действий и предвидения их результата получены при сравнении орудийного поведения шимпанзе и павианов.
Отрицательные результаты, полученные в опытах с низшими обезьянами, не позволяют выяснить их причину: животное или не способно оценить умственное состояние другой особи, или, сделав такую оценку, оно не может ее использовать.
Продолжая сравнение низших обезьян с шимпанзе, следует упомянуть эксперименты Примэка с шимпанзе Сарой, в которых обезьяна должна была отличить состояние знания и незнания у человека. Ей показывали видеозаписи действий дрессировщиков, пытающихся решить различные бытовые проблемы, например включить кнопкой магнитофон, у которого шнур выдернут из розетки. Во всех случаях Сара указывала возможное верное решение для любимого дрессировщика и ... неверное — для нелюбимого. Эти данные доказывают способность шимпанзе поставить себя на место другого и мысленно воспроизвести его действия, а также свидетельствуют о его способности к осознанному обману. Ряд наблюдателей приводят примеры того, как шимпанзе обманывают друг друга в различных ситуациях с помощью разнообразных жестов, поз, мимики.
5.4. Коммуникация и язык животных
Общественная организация животных в целом представляет собой суммарный результат взаимодействия между членами сообщества.
Коммуникация составляет сущность любого социального поведения. Трудно представить себе общественное поведение без обмена информацией или же систему передачи информации, которая не была бы в каком-то смысле общественной. Когда животное совершает некое действие, изменяющее поведение другой особи, можно говорить о том, что имела место передача информации. Это весьма широкое определение, под которое подходят и те случаи, когда, например, спокойно кормящееся или, наоборот, тревожно насторожившееся животное только лишь своей позой воздействует на поведение других членов сообщества. Поэтому этологи, изучающие процесс коммуникации, задаются вопросом: "намеренно" ли передается сигнал или он лишь отражает физиологическое и эмоциональное состояние животного? Могут ли такие социальные животные, как обезьяны, дельфины, волки или муравьи передать друг другу точные сведения, например о том, в какой точке пространства находится источник пищи и как этой точки удобнее достичь. Исследование пределов коммуникативных возможностей животных — одна из самых интересных и дискуссионных проблем в этологии.
5.4.1. КАК РАБОТАЮТ СИГНАЛЫ
Известно, что разные группы животных более или менее специализированы по сенсорной модальности используемых сигналов, в зависимости от степени развития у них тех или иных органов чувств. Так, тактильная коммуникация доминирует во взаимодействиях многих беспозвоночных, например у слепых рабочих термитов, которые никогда не покидают своих подземных туннелей, или у дождевых червей, которые ночью выползают из нор для спаривания. У беспозвоночных осязание тесно связано с химической чувствительностью, так как специализированные тактильные органы, например антенны насекомых, часто снабжены хеморецепторами. Общественные насекомые передают большое количество информации путем комбинации тактильных и химических сигналов.
Тактильная коммуникация в силу своей природы возможна только на близком расстоянии. Длинные антенны тараканов и раков позволяют им исследовать мир в радиусе одной длины тела, но это почти предел для осязания. Тактильная коммуникация сохраняет свое значение у многих позвоночных, в частности у млекопитающих, наиболее "общественные" виды которых проводят значительную часть времени в физическом контакте друг с другом. У приматов важнейшую форму социальных контактов составляет обыскивание шерсти, или "груминг".
Другие сенсорные системы — системы зрения, слуха и обоняния обеспечивают общение на значительном расстоянии. Звук и запах имеют дополнительное преимущество благодаря тому, что они способны преодолевать естественные препятствия, например густую растительность. Звуковые сигналы. Сигналами, рассчитанными на большое расстояние, являются обычно крики. Например, территориальные птицы поют свои песни, выбирая самую высокую точку местности ("песенный пост"). Птицы открытых ландшафтов (жаворонки, луговые коньки) поют, летая высоко над своей территорией. Рекорд дальности в звуковой коммуникации животных поставлен горбатыми китами: их песни могут восприниматься другими китами, находящимися на расстоянии нескольких десятков километров.
Химические сигналы особенно хорошо развиты у насекомых и у млекопитающих. Феромоны бабочек улавливаются самцами с подветренной стороны за 4—5 км, и они самые устойчивые из феромонов насекомых. Наоборот, феромоны тревоги, используемые муравьями, распространяются на расстояние в несколько сантиметров и быстро распадаются, что вполне объяснимо, так как эти сигналы призваны обеспечить быструю, но локальную реакцию. Многие млекопитающие маркируют свою территорию при помощи запаховых меток, часто концентрируя их на особых местах. Запах используется также для сообщений о состоянии самок млекопитающих в период размножения. Во время эструса они выделяют специальные химические вещества, сигнализируя этим готовность к встрече с самцом. Этот тип сигнала используется у самых разных видов. Он аналогичен, например, запахам, издаваемым неоплодотворенными самками бабочек и привлекающим самцов с весьма далеких расстояний. Такие вещества называются половыми феромонами (см. также: 5.2.3).
Зрительные сигналы могут действовать лишь на относительно коротком, в пределах видимости, расстоянии. Исключение составляют простые сигналы тревоги, в виде белых пятен на теле, например хвосты оленей и кроликов, видимые на большом расстоянии. К зрительным сигналам относятся также широко распространенные опознавательные знаки, многие из которых действуют по принципу "есть или нет". Так, ключевым признаком для определения пола у американского золотистого дятла является наличие черных усиков у самца, и если вклеить их самке, как это сделал Г.Нобль, самец сразу прогоняет ее (рис. 5.17). Ритуальные демонстрации и танцы брачных партнеров также воздействуют в первую очередь на зрительный анализатор (рис. 5.18).

Рис. 5.17. Самец (слева) и самка (справа) золотистого дятла. Ключевой признак для определения пола птицами своего вида — черные усики самца

Рис. 5. 18. Совместная брачная демонстрация у манакина. Часть демонстрации заключается в "кувыркании" двух самцов перед самкой (по Alcock, 1989)
О. Меннинг отмечает связь частоты использования цветовых зрительных сигналов с развитием цветного зрения у животных. Сложная брачная демонстрация бриллиантовой амадины включает в себя и "преподнесение подарка" (рис. 5.19). Оно практически универсально для многих групп, за исключением большинства млекопитающих. В ранней истории млекопитающих, когда их предки были ночными насекомоядными, цветовое зрение давало им мало преимуществ и почти утратило роль в процессе коммуникации. Поэтому яркая разноцветная раскраска некоторых рыб, рептилий и птиц поразительно контрастирует с универсальной серой, черной и коричневой окраской млекопитающих. Исключение составляют некоторые белки и приматы — это как раз те две группы млекопитающих, у которых цветовое зрение более развито, чем у остальных. В их окраске встречаются синие, красные, зеленые и желтые участки, которые они могут демонстрировать, например, в ритуалах угроз или подчинения.

Рис.5.19. Ухаживание у бриллиантовой амадины Steganopleura guttata. Самец демонстрирует самке подарок — травинку (а), поет и выполняет танец (б), а затем, сохраняя травинку в клюве, принимает позу выпрашивания пищи, характерную для молодых птиц (в) (по Eibl-Eibesfeldt, 1977)
В естественных условиях сигналы различных модальностей часто объединены в эффективные комбинации, сочетающие, например, и звук, и зрительный стимул. Хорошим примером являются брачные ритуалы райских птиц, включающие характерные позы, демонстрации "ритуальных" перьев, прыжки, крики и хлопанье крыльями.
Один из примеров такой комбинации, также действующей в сфере полового поведения, — строительство и украшение шалашей, которые служат местом интенсивного соперничества между самцами у 14 из 18 видов шалашников, или беседковых птиц (сем. Ptilonorhynchidae), населяющих Австралию и Новую Гвинею. У двух видов — это беседка, достигающая полутораметровой высоты, в центре которой находится разукрашенное птицей деревце, а сводчатый коридор ведет из беседки на расчищенную площадку для тока. Для украшения беседки используются перья, раковины, кусочки костей, надкрылья жуков и цикад, а возле поселений человека еще и монеты, булавки, стекло, бижутерия, чайные ложечки, гвоздики, винтики, бумага и тому подобные предметы, сочетающиеся по цвету с украшениями природного происхождения (рис. 5.20). У некоторых видов самцы к тому же раскрашивают шалаши, смешивая сок ягод со слюной. Это пример, по-видимому, одного из самых сложных зрительных сигналов (см. также: Резникова, 1997).

Рис. 5.20. Самец австралийского атласного шалашника у своего шалаша (рис. О. Перепелкиной)
Таким образом, сигналы, используемые животными, весьма многочисленны. Однако все их многообразие у разных видов укладывается приблизительно в 10 основных категорий:
1) сигналы, предназначенные половым партнерам и возможным половым конкурентам;
2) сигналы, которые обеспечивают обмен информацией между родителями и потомством;
3) крики тревоги, зачастую воспринимаемые животными разных видов;
4) сообщения о наличии пищи;
5) сигналы, помогающие поддерживать контакт между общественными животными, например перекличка шакалов или крики стайных птиц; Лоренц (1984) описывает своеобразное "голосование" у стаи гусей: сначала одна птица издает "стайный крик", и если большинство птиц его подхватывает, стая взлетает, но если ответило слишком мало птиц, то начавшая кричать птица замолкает и стая остается на земле;
6) сигналы - "переключатели", предназначенные подготовить животное к действию последующих стимулов. Явление, связанное с такими сигналами, названо метакоммуникацией. Так, поза с прижатыми к земле передними лапами предшествует драке-игре у львов и у псовых, она не встречается ни в каких других ситуациях и говорит о том, что все последующие агрессивные действия — это всего лишь игра (см.: Зорина, 1998). У обезьян в таких случаях появляются особая, "игровая" мимика и походка (Гудолл, 1992);
7) сигналы-"намерения", которые предшествуют совершению какой-то реакции: например, перед взлетом птицы совершают особые движения крыльями;
8) сигналы, связанные с выражением агрессии;
9) сигналы миролюбия;
10) сигналы фрустрации.
До сих пор мы рассматривали работу различных сигналов, не обсуждая вопроса о наличии четких границ, разделяющих отдельные единицы коммуникации. Ведут ли животные диалог по принципу "вопрос-ответ", или сигналы плавно переходят один в другой, образуя континуум, в котором партнеры могут выделять отдельные фрагменты и реагировать на них? Этологи высказывали представление о том, что одни сигналы могут действовать как триггеры ("спусковые крючки"), а другие — как "насосы". Крики тревоги, например, постоянны для каждого вида. Они действуют как триггеры, резко меняя поведение особи. О принципе "насоса" говорят тогда, когда влияния сигналов постепенно накапливаются и изменяют вероятность того, что получатель будет отвечать. Например, самцы голубей при ухаживании в течение нескольких дней много раз повторяют характерные "поклоны", прежде чем произведут на самку нужное впечатление. В этом случае градуированное воздействие заключается в накоплении эффекта от повторения. Наряду с этим есть сигналы, которые сами по себе могут быть градуированными. Так, при агрессивных стычках животные нередко испускают различные крики или осуществляют зрительные демонстрации (движения хвоста у собак, приподнимание хохолка у соек). Эти сигналы имеют несколько градаций, которые коррелируют с вероятностью того, что животное нападет или убежит. Приведенный выше рис. 5.6, на котором показаны соответствующие "градации" в мимике кошки (по Лейхаузену), представляет собой иллюстрацию этого положения.
В целом создается впечатление, что коммуникационные возможности большинства видов животных весьма специализированы и ограничены, и создание новых сигналов может происходить при их комбинировании или градуировании. Например, М. Килей (Кillеу, 1972), подробно исследуя звуковые сигналы копытных животных, пришла к выводу о том, что большинство их континуально (непрерывно) и выражает уровни возбуждения и основные мотивационные состояния животного. Она считает, что специфических звуков, соответствующих определенным состояниям ("слов"), у копытных ничтожно мало, и даже крайние элементы континуумов можно связать переходными формами. К такому же выводу приходит Е.Н. Панов (1978, 1983) на основе анализа системы коммуникации многих видов животных. В то же время этологи, исследующие сообщества высокосоциальных и психически высокоорганизованных видов, сталкиваются с пограничными ситуациями, когда возникает предположение, что в некоторых случаях их система коммуникации выходит за указанные рамки (см.: 5.4.4.5). Проблеме коммуникации у животных посвящено огромное число исследований. Рассмотрим лишь некоторые из них.
5.4.2. ЯЗЫК ЖИВОТНЫХ И МЕТОДЫ ЕГО ИЗУЧЕНИЯ
Долгое время абсолютное большинство исследователей считали, что наличие языка — это уникальное свойство человека. Поскольку существует тенденция определять язык таким образом, что он предстает исключительной привилегией людей, это вносит в исследования определенные предубеждения. Существует так много определений языка, что только их перечислению посвящена первая глава книги известного американского психолингвиста Н. Хомски "Язык и сознание" (Chomsky, 1980).
Как отмечает в книге "Поведение животных" Д. Мак-Фарленд (1982), определить понятие языка с объективной точки зрения нелегко, поскольку он характеризуется многими необходимыми признаками. Например, мы можем согласиться с тем, что язык — это средство коммуникации, но очевидно, что не все средства коммуникации являются языком. Язык человека обычно существует в форме речи, но это далеко не всегда так (азбука Морзе, языки там-тамов, сигналы дымных костров и семафоров, "свистовые" языки некоторых народов обычно рассматриваются как вспомогательные языковые средства). Вместе с тем, символический язык, используемый при общении глухонемых, — это грамматически организованная знаковая система. Язык человека использует символы, но также символичны и некоторые аспекты коммуникации у пчел. Люди осваивают язык в течение специфического чувствительного периода развития, но то же самое наблюдается у некоторых птиц, когда они учатся песне своего вида.
С помощью языка можно передавать информацию не только о сиюминутных ситуациях, но и о таких, которые удалены и во времени, и в пространстве (свойство перемещаемости, см.: 5.4.4.3). Однако некоторые сигналы тревоги у животных обладают теми же свойствами.
Использование человеком грамматических правил — это тот аспект языка, который, казалось бы, явно выделяет его из коммуникативных систем животных. Тем не менее, благодаря некоторым исследованиям на шимпанзе (Savage-Rumbaugh et al., 1993) и это положение перестает быть бесспорным. Несомненно, язык человека гораздо сложнее языка животных. Но означает ли это, что между общением людей и общением животных существует качественная разница, или все дело в степени развития этого общения? Многие исследователи пытались ответить на этот непростой вопрос, применяя принципиально различные подходы и методы.
5.4.3. ПОПЫТКИ ПРЯМОЙ РАСШИФРОВКИ ЯЗЫКА ЖИВОТНЫХ
5.4.3.1. ТАНЦЫ ПЧЕЛ.
Одним из самых выдающихся достижений в этой области, видимо, следует считать открытие и подробное исследование символического "языка танцев" медоносной пчелы Карлом фон Фришем (Frisch, 1923; Фриш, 1980), получившим за эти исследования Нобелевскую премию в 1973 г. (тогда же ее получили и два других выдающихся этолога: К. Лоренц и Н. Тинберген). Сам факт способности пчел передавать информацию о месте массового цветения и наличия у пчел-разведчиц стереотипных повторяющихся движений зафиксирован еще в 1788 г. Шпитцнером, однако Фриш впервые исследовал это явление именно как язык (рис. 5.21). Он показал, что пчелы используют разные по степени конкретности способы дистанционного наведения: простой круговой танец передает информацию о том, что нужно "искать в пределах 100 м", и стимулирует рабочих покинуть улей. Если же необходимо мобилизовать пчел на дальний поиск, разведчица исполняет внутри улья, на сотах, виляющий танец. В нем угол, образуемый между прямой пробега танцовщицы на сотах и направлением силы тяжести, сигнализирует о направлении полета (причем пчелы-наблюдательницы переводят значение угла танца по отношению к силе тяжести снова в угол по отношению к Солнцу). Расстояние до источника корма коррелирует с 11 параметрами танца, например с его продолжительностью, темпом, количеством виляний брюшком, с длительностью звуковых сигналов.
Рис. 5.21. Танец медоносной пчелы.
Таким образом, по выражению О. Меннинга (1982), "мир вынужден был признать, что передавать информацию в символической форме может не только человек — это способно сделать такое скромное создание, как пчела". Однако открытие Фриша было признано отнюдь не безоговорочно. В 60— 70-е годы разгорелась бурная дискуссия по поводу того, действительно ли пчелы передают информацию с помощью системы дистанционного наведения, включающей абстрактные символы, или они в какой-либо форме используют пахучие вещества. Было высказано предположение о том, что идеальным разрешением этого спора были бы результаты, полученные с помощью пчелы-робота — модели, изготовленной для выполнения танца под контролем человека. Первая удачная попытка такого рода была осуществлена Н.Г. Лопатиной (1971): ей удалось вступить в диалог с семьей пчел, которые воспринимали "танец" модели пчелы-танцовщицы с информацией о расстоянии до кормушек. Позднее более детальные и весьма эффективные опыты с искусственными пчелами были проделаны датским исследователем А. Михельсеном (рис. 5.22; Michelsen et al., 1990).
Рис. 5.22. Модель "танцующей" пчелы, предложенная Михельсеном (Michelsen et al., 1988). а — схема модели, б — использование модели в улье. "Пчела" состоит из тела (1) и крыла (2), сделанного из куска лезвия бритвы. К области головы модели через фитилек (3) с помощью трубки (4) подается сахарный раствор, который слизывают пчелы-наблюдатели. Специальные устройства обеспечивают генерацию и передачу вибраций на "крыло" (5, 6) и движения "тела", как при естественном танце (7)
Так был подтвержден сам факт использования пчелами "языка танцев". Расшифровка этого языка до сих пор не завершена. Новые исследования в этой области приносят интересные результаты, но они же ставят и новые вопросы. Е.К. Еськов (1979) экспериментально исследовал звуковую составляющую танца пчел и установил, что акустический сигнал дальности может нести информацию о расстоянии до самых различных целей полета (источники углеводной и белковой пищи, новое жилище), причем пчелы способны корректировать структуру своего акустического сигнала в зависимости от структуры акустических помех. Один из последователей Фриша М. Линдауэр показал, что пчелы используют визуальные компоненты танца для передачи сведений не только о пище, но и для указания подходящего места для жилья во время роения.
Были выделены также новые, ранее неизвестные параметры и компоненты танца. Стали известны "отпугивающие" танцы при угрозе отравления инсектицидами. Большой вклад в исследования коммуникации пчел внес И.А. Левченко (1976). Он сопровождал киносъемкой эксперименты, в которых пчелы решали различные пространственные задачи с помощью коммуникации, и анализировал геометрические фигуры танца и различные составляющие передаваемых сигналов. Оказалось, что все формы фигур танцев включают одни и те же элементы и различаются количеством "виляний" в прямолинейном пробеге. Степень точности целеуказания прямо коррелирует с расстоянием до цели. Количество звуковых сигналов, издаваемых разведчицей во время танца, также зависит от расстояния до цели. При этом информацию воспринимают не все пчелы в свите разведчицы, а лишь те, которые следовали за ней на протяжении 4—9 циклов танца.
О том, что возможности пчелиного языка еще далеко не познаны, говорят и опыты самого К. Фриша (1980), в которых танцовщицы вынуждены были огибать холм, но в танце они указывали направление по прямой, а расстояние — с учетом усилий, затраченных при полете вокруг холма. По выражению автора, это относится "...к наиболее удивительному чуду, которыми так богата жизнь пчел". Эту ситуацию позднее формализовал Бизецкий: он помещал перед ульем туннель, в конце которого находилась чашечка с сиропом. Бели туннель был прямой, то танец указывал правильное направление; если туннель делал поворот под прямым углом, то направление, указанное пчелами, соответствовало гипотенузе угла, а расстояние было близким к истинному. Если же туннелю придавали круглую или V-образную форму, то танцы были очень сложными и расшифровать их не удавалось.
Вместе с тем, есть примеры, свидетельствующие об ограниченности языка пчел. Так, Фриш отметил, что в нем нет слова "вверх" ("...цветы не растут в небесах"), и пчелы могут передавать информацию о перемещении кормушки только в горизонтальной плоскости. Он установил это в опыте, когда улей находился внизу радиобашни, а кормушка наверху. Ее показали пчелам-разведчицам, но они не смогли мобилизовать сборщиц. По мнению многих современных этологов и психолингвистов, это свидетельствует о "закрытости" языка танцев, тогда как языки человека являются открытой системой. Обладая так называемым свойством "продуктивности" (см.: 5.4.4.3), они способны создавать неограниченное количество сообщений о чем угодно. Заметим, однако, что если бы гипотетические наблюдатели предложили человеку нечто соблазнительное, поместив его в пятое (или ...энное) измерение, в нашем языке могло бы и не хватить средств для передачи такой информации.
Еще один, ставший классическим, результат расшифровки естественных сигналов животных был получен Т.Струзейкером (Strusaker, 1977), описавшим символические акустические сигналы у восточноафриканских верветок (Cercopithecus aethiops). Обезьяны издавали по-разному звучащие крики в ответ на появление трех хищников: леопардов, орлов и змей. При этом сигналы, издаваемые при появлении леопарда, заставляли верветок взбираться на деревья, тревога по поводу орла — вглядываться в небо и спасаться в кустах, а при звуках, означающих появление змеи, обезьяны становились на задние лапы и вглядывались в траву. Таким образом, впервые было точно показано, что они используют различные знаки для маркировки разных предметов или разных видов опасности.
Существовали, однако, и более скептические интерпретации: тревожные крики могут служить просто сигналами общей готовности, заставляющими животных оглядываться, и если они видят хищника, то реагируют на увиденное, а не на услышанное. Возможно и то, что крики обезьян — это не символы разных хищников, а выражение относительной интенсивности испуга, вызываемого леопардами, орлами и змеями. В этом случае возможность считать сигналы верветок семантическими становилась еще менее вероятной.
Эти гипотезы были проверены Сифартом и Чини (1993; Seyfarth et al., 1980). Они транслировали обезьянам записи различных сигналов, естественно, в отсутствие хищников, так что животные реагировали только на звуковую, а не на зрительную составляющую этого сложного комплекса стимулов. Чтобы проверить, не отражают ли сигналы животных степень испуга или возбуждения, записи изменяли, делая их длиннее или короче, громче или тише. Результаты свидетельствовали о том, что крики тревоги выполняли функцию именно семантических сигналов.
В исследовании аналогий между сигналами обезьян и словами нашего языка авторы пошли дальше. Известно, что при сравнении слов мы делаем заключение об их сходстве или различии на основе их смысла, а не акустических свойств. Так, мы считаем слова "предательство" и "измена" сходными, а слова "предательство" и "обстоятельство" — различными. Опыты четко показали, что верветки оценивают различные звуки именно на основе их смыслового содержания, а не только акустических свойств.
Семантическая система обезьян этого вида, видимо, несложная: некоторые крики (например, предупреждающие о появлении леопарда и змеи) различаются по смыслу, а другие (например, "врр-р" и щебет) относятся к одному и тому же явлению и могут выражать разные оттенки в пределах одной понятийной категории.
Данные Сифарта и Чини интересно сопоставить с наблюдениями за другими животными. Дж. Гудолл (1992. С. 146—148) не проводила специальных исследований этого вопроса, но отмечала, что некоторые издаваемые шимпанзе звуки могут различаться в зависимости от вида пищи или конкретной опасности. Например, звук "хуу" издается только при виде небольшой змеи, неизвестного шевелящегося создания или мертвого животного.
Проанализированные случаи успешной "расшифровки" семантических сигналов можно объяснить на редкость удачно выбранными объектами, хотя и столь несходными между собой: речь идет о сравнительно четко выраженных отдельных сигналах (фигуры пчелиного танца, крики верветок), которые соответствуют четким ситуациям (поиск точки в пространстве, появление орла в небе или змеи в траве). Такие ситуации могут служить ключом для расшифровки "речи" животных. В большинстве же случаев определить смысл, например, сложнейших акустических сигналов дельфинов или волков, а также быстрых движений "языка жестов" муравьев достаточно трудно. Такие трудности сопоставимы с ситуацией лингвиста, у которого есть отрывки рукописей на неизвестном языке и нет ключа к ним.
В эксперименте можно заставить животное решать определенную задачу, для которой необходимо использование коммуникации, т.е. можно смоделировать подобную ситуацию, однако сами сигналы либо не поддаются фиксации (примером может служить описанный ниже эксперимент Э.Мензела с шимпанзе), либо крайне неудобны для классификации. Представим, что мы наблюдаем действия японцев во время чайной церемонии и пытаемся по произносимым словам и совершаемым действиям составить русско-японский словарь. При этом наблюдатель не понимает смысла большинства действий, не знает, где начинаются и кончаются слога и фразы, одно и то же слово, произносимое разными лицами, может воспринимать как разные слова и, наоборот, несколько слов принимать за одно. По-видимому, из таких наблюдений составить словарь невозможно.
Исследователь акустической коммуникации волков (Теберг) отмечал удивительную способность волка к тончайшему различению ничтожных оттенков звука и считал, что она не может быть просто игрой природы. Автору удалось вычленить несколько постоянных сигналов (например, "звук одиночества", когда один из членов стаи попал в изоляцию и стремится присоединиться к остальным), а также выяснить, что волки могут передавать какую-то конкретную информацию небольшим изменением отдельных параметров голоса (см. также: 5.5.2).
Исследования акустической коммуникации млекопитающих принесли интересные результаты, однако до сих пор естественные семантические системы даже таких высокоорганизованных социальных животных, как шимпанзе или дельфины не могут считаться расшифрованными. Ниже мы рассмотрим два примера, связанные с этими животными, когда четко поставленные эксперименты свидетельствуют о передаче сложной информации, но сам язык животных не поддается фиксации или расшифровке.
Многочисленные попытки предпринимались для расшифровки языка дельфинов (см., например: Марков, 1993). Один из первых и наиболее знаменитых экспериментов был проведен В. Эвансом и Дж. Бастианом (Evans, Bastian, 1969) с парой дельфинов, которые должны были сообщить один другому, в каком порядке нужно нажимать на педали, чтобы получить рыбу. Первоначально обоих дельфинов содержали вместе, и каждый из них научился нажимать левую педаль, если светящаяся лампочка начинала мигать, и правую, если она светилась стабильно. Затем их помещали в разгороженный пополам бассейн так, что они не могли видеть друг друга и общались лишь с помощью акустических сигналов. Лампочка была видна только одному из дельфинов, а педали были в обоих отсеках бассейна. Животные получали вознаграждение в том случае, когда оба нажимали на педали правильно. Успех, достигнутый дельфинами в тысячах испытаний, говорит об их способности передавать конкретную информацию.
Аналогичные опыты были проведены В.И. Марковым и сотр. с использованием модифицированной методики (см.: Zanin et al., 1990). Они подтвердили, что афалины могут координировать поведение друг друга во время опыта и передавать информацию либо о размере мяча (маленький — большой), либо о том, в каком положении его предъявляют (справа — слева).
Трактовка этих экспериментов до сих пор вызывает споры, и приведенные данные свидетельствуют о значительных препятствиях на пути расшифровки естественных коммуникативных систем. Американский исследователь Э. Мензел (Menzel, 1979) провел серию экспериментов, в которых выяснял, могут ли шимпанзе передавать друг другу информацию о местоположении пищи, не прибегая к прямому показу, а используя "дистанционное наведение". В сопровождении одной из обезьян он прятал пищу, а затем выпускал всех шестерых шимпанзе, предоставляя им возможность найти тайник. Специально фиксировались только те, довольно многочисленные, случаи, когда обезьяна-свидетель не возглавляла группу, а двигалась сбоку или сзади, но, тем не менее, группа бежала прямо к тайнику. Когда Мензел вместо пищи прятал "страшную" игрушку (например, змею), шимпанзе приближались к тайнику осторожно, с явными признаками страха. В отдельных сериях тестов двум шимпанзе показывали разные тайники. Когда всех обезьян после воссоединения со "свидетелями" выпускали, они выбирали из этих тайников наиболее привлекательный: либо предпочитали большее количество пищи меньшему, либо фрукты — овощам. Несмотря на столь красноречивые результаты, Мензелу не удалось установить, с помощью каких конкретных сигналов передается эта информация. По его мнению, речь может идти о тонких мимических движениях, которых наши средства наблюдения не улавливают (подробнее см.: Панов, 1983).
О том, что шимпанзе обладают потенциальной способностью к передаче не только видоспецифических сигналов, но и совершенно новой информации, свидетельствуют опыты по обучению обезьян языкам-посредникам (см.: 5.4.4).
Выше (см.: 2.7.4) мы кратко описали историю этого поистине революционного направления в изучении поведения, получившего название "language-training experiments". Оно составило важный этап в развитии и этологии, и физиологии высшей нервной деятельности, и, наконец, психолингвистики. Обучение обезьян искусственным языкам (так называемым языкам-посредникам), которые в той или иной степени обладают свойствами человеческой речи, сыграло и продолжает играть важнейшую роль в двух неразрывно связанных, но все же самостоятельных аспектах изучения поведения. Первый касается оценки уровня развития когнитивных процессов — способности к образованию довербальных понятий и использованию символов вместо реальных предметов и явлений (Premack, 1983; 1984; Rumbaugh et al., 1984; 1991; Gardner, Gardner, 1985). В настоящей главе рассматривается второй аспект и делается попытка описать поведение обезьян в процессе обучения языкам-посредникам как проявление наиболее сложной формы коммуникации у животных (см. табл. 5.2). Исследование этих вопросов важно для понимания эволюции поведения, так как позволяет оценить, какие элементы коммуникативных процессов животных и в какой степени предшествовали появлению речи человека.
Наука шла к этому эксперименту с первых шагов исследования высших форм психики животных, в первую очередь шимпанзе. Этих человекообразных обезьян недаром называют лабораторным двойником человека. Действительно, среди современных животных это наши ближайшие родственники: у человека и шимпанзе очень много общих генов, а также много общего в строении и функционировании всех органов, включая мозг. Более того, по многим особенностям своей психики шимпанзе гораздо больше похожи на человека, чем на других животных, включая и приматов.
Тем не менее долгое время считалось, что имеется одна ступень в развитии психики, на которую человекообразные обезьяны не смогли не только подняться, но даже к ней приблизиться (настолько принципиальными считались особенности, лежащие в ее основе). Эта ступень — человеческая речь, которая служит не только средством общения, но и аппаратом мышления. Действительно, как мы уже убедились, язык животных, включая и обезьян, — это конкретные звуковые, обонятельные, зрительные и т.п. сигналы, которые действуют в данной ситуации, в данный конкретный момент. Они одинаковы у всех особей каждого вида, запас их определен генотипически и практически не подлежит расширению. И.П. Павлов называл их "первой сигнальной системой", общей для животных и человека. В отличие от этого язык человека позволяет передавать информацию в отвлеченной форме с помощью слов-символов, которые являются сигналами других, конкретных сигналов. Именно поэтому И.П. Павлов называл слово "сигналом сигналов", а речь — "второй сигнальной системой", которая позволяет не только реагировать на конкретные стимулы и сиюминутные события, но в отвлеченной форме передавать информацию об отсутствующих предметах, о событиях прошлого и будущего.
Долгое время существовало, да и сейчас еще не полностью изжито представление, что между психикой человека и его близких и далеких родственников лежит непроходимая пропасть, так что речь человека не имеет никаких биологических корней. Такая точка зрения не была единственной, но именно она господствовала в официальной советской физиологии 50-х годов. Среди современных ученых, в том числе и западных, также до сих пор имеются активные сторонники представления о том, что никаких элементов этой формы психики у антропоидов нет и быть не может (Terrace et al., 1979, 1985).
Однако чем больше появлялось экспериментальных данных о сложности высшей нервной деятельности человекообразных обезьян, тем более логичным становилось предположение о том, что наряду с развитой первой сигнальной системой они обладают и какими-то, пусть самыми примитивными зачатками второй сигнальной системы. Как мы уже показали в разделе 2.7.4, вопрос о том, в какой мере и форме у антропоидов может проявляться способность пользоваться символами вместо реальных предметов и явлений, закономерно возникал у многих исследователей, и, начиная с 70-х годов, анализом этой проблемы прицельно занимается несколько научных коллективов. Супруги Аллен и Беатрис Гарднер (Gardner, Gardner, 1969; 1985; см. также: 2.7.4), выбрав жестовый язык американских глухих — амслен (AMerican Sign LANguage), получили возможность исследовать способности шимпанзе овладевать элементами языка, построенного по правилам английской грамматики. Их подход и метод были вполне адекватны для того, чтобы выяснить, в какой мере такой язык может стать средством коммуникации обезьяны и человека, а также обезьян между собой. Первая воспитанница Гарднеров — шимпанзе Уошо — за 3 года обучения усвоила 130 знаков, изображенных сложенными определенным образом пальцами (рис. 5.23).

Рис. 5.23. Примеры знаков амслена, усвоенных Уошо: "сладость и конфета"; "пить"; "веревка"; "книга" (По Линдену, 19S1)
Далее по собственной инициативе она начала их комбинировать в цепочки из 2—5 слов. Первыми такими комбинациями были "дай сладкий" и "подойди открой". Позднее Гарднеры работали и с другими шимпанзе. Эти и другие опыты подробно и достоверно описаны в книге известного американского популяризатора Ю. Линдена (1981; см. также: Ерахтин, Портнов, 1984; Мак-Фарленд, 1988).
Все "разговоры" Уошо и части других обезьян фиксировались на видеопленку. Часть экспериментов всех перечисленных авторов была организована по методу двойного слепого контроля: половина экспериментаторов не знала ответа на вопрос, предлагаемый обезьяне: она должна была называть объекты, изображенные на слайде, делая соответствующий знак тренеру, который, в свою очередь, не видел этого слайда. Второй экспериментатор, которого обезьяна не видела, фиксировал ее жесты, но не знал, какие слайды ей демонстрируются. В этой ситуации Уошо правильно ответила на 92 из 128 вопросов.
Специальные опыты показали, что шимпанзе не просто вырабатывают ассоциации между знаками и соответствующими предметами и действиями, но понимают их смысл. Ряд особенностей, проявившихся при пользовании обезьян своим словарным запасом, свидетельствовал, что запоминание знаков сопровождается процессами более высокого порядка — обобщением и абстрагированием. В частности, оказалось, что, несмотря на отсутствие специальной процедуры, обезьяны применяют усвоенные жесты к довольно широкому набору незнакомых предметов той же категории, что и использованные при обучении. Так, например, шимпанзе Ним в опытах Герберта Терреса (Terrace et al., 1979), как и все остальные обезьяны, делал соответствующий знак, когда видел представителей любых пород собак, не только живых, но и на картинках, или если просто слышал собачий лай. Иными словами, обезьяны переносят навык называния предмета с единичного образца, использованного при обучении, на все предметы данной категории.
Усвоенную систему знаков шимпанзе использовали как средство классификации предметов и их свойств. Впервые это было четко показано Роджером Футсом (Fouts, 1975) в опытах на шимпанзе Люси, которая имела относительно ограниченный запас знаков (60), но с их помощью почти безошибочно относила к соответствующей категории новые, ранее никогда ей не предъявлявшиеся овощи, фрукты, предметы обихода и т.п. Обезьянам оказались доступны переносы значений символа, иногда довольно тонкие. Так, Уошо назвала служителя, долго не дававшего ей пить, "грязный Джек", и это слово явно было употреблено не в смысле "запачканный", а как ругательство. В других случаях разные шимпанзе называли бродячего кота "грязным котом", а гиббонов — "грязными обезьянами". Люси применяла для обозначения невкусного редиса знаки "боль" и "плакать".
Чтобы обучить Уошо знаку "нет", Гарднеры просигналили ей, что снаружи ходит большая собака, которая хочет ее съесть. Через некоторое время обезьяне предложили погулять, и она отказалась. Единственной причиной могло быть воспоминание о собаке, причем образ собаки приобрел дополнительный признак — "быть снаружи". Он стал посредником между образами "гулять" и "собака". Последующие эксперименты по той же методике, проведенные на других обезьянах (Patterson, 1978), показали, что словарь даже в 400 знаков далеко не исчерпывает их возможностей. Кроме того, как отмечали Гарднеры (1985), в болыдинстве случаев опыты проводились на молодых шимпанзе и прекращались, самое позднее, когда им исполнялось 10 лет. Учитывая, что в неволе они могут жить до 50 лет, авторы допускали, что полученные данные отражают далеко не все возможности этих животных.
Еще один важный аспект проблемы — возраст, когда начинается обучение. Опыты на Уошо, одни из наиболее успешных, начались, когда ей было 10 месяцев. Сравнение языковых навыков разных шимпанзе (Лана, Шерман и Остин, Кэнзи), исследованных, начиная с 1972 г., Д. Рамбо и С. Сэведж-Рамбо, также показывает, чем раньше начато обучение, тем больших результатов достигают обезьяны. Так, карликовый шимпанзе Кэнзи, который с 10 месяцев постоянно слышал разнообразную человеческую речь (в соответствии со специальной программой), в возрасте 5 лет спонтанно обнаружил способность понимать ее в существенно большем объеме, чем все остальные обезьяны (Savage-Rumbaugh et al., 1993).
Сам по себе факт заучивания знаков еще не несет ничего принципиально нового — для этого достаточно простого условнорефлекторного обучения. В таком случае феномен легко можно было бы объяснить в рамках бихевиористских представлений, как это делали и до сих пор продолжают делать многие авторы (например, Schusterman, Gisiner, 1988). Тем не менее ряд особенностей пользования "словарным запасом" заставлял предполагать, что запоминание знаков сопровождается у шимпанзе процессами более высокого порядка, включающими формирование обобщений, в том числе высокоабстрактных. Об этом свидетельствует тот факт, что Уошо, а затем и другие обезьяны были способны запоминать не только знаки (жесты, пластиковые жетоны, компьютерные символы и т.п.), обозначающие окружающие предметы, их свойства, основные действия, совершаемые шимпанзе и человеком, но также некоторые отвлеченные понятия типа "еще", "нет", эмоциональные состояния — "больно", "смешно" и т.п. Они могли использовать знаки в переносном смысле и в новых ситуациях, образовывать из них 2—3-членные предложения с соблюдением порядка слов, характерного для английского языка (табл. 5.2).
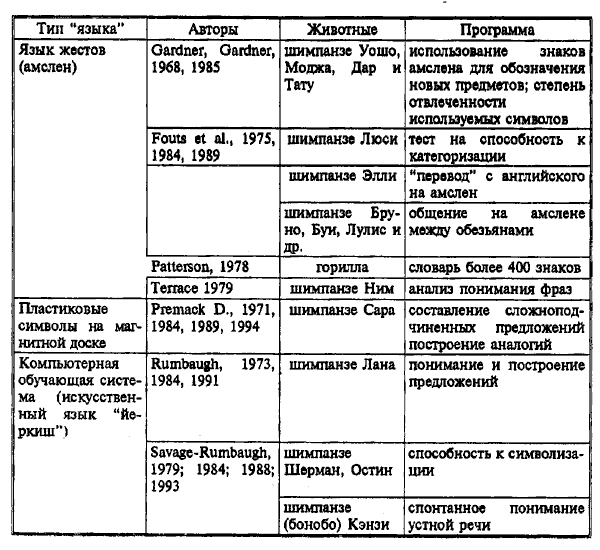
Таблица 5-2. Основные программы обучения обезьян "«зыкам-посреднюим"
Наряду с шимпанзе языку жестов обучали также гориллу (Patterson, 1978) и орангутанов (Miles, 1983). Кроме того, ряд ученых попытались исследовать эту форму коммуникации у животных-неприматов.
Способность морских млекопитающих к овладению языками-посредниками в течение ряда лет изучает американский исследователь Л. Херман (Herman, 1986). В его работах дельфины афалины должны были сначала усвоить "названия" различных предметов, находящихся в бассейне, и совершаемых с ними действий. "Словами" служили жестовые сигналы экспериментатора, который стоял на краю бассейна. После того как дельфины устанавливали связи между жестами и объектами в бассейне, а также между жестами и манипуляциями, которые они должны были выполнить, проводились тесты с использованием новых предложений, в которых их просили принести или переместить какой-либо предмет, либо поместить один предмет внутрь другого или на него, или под него и т.п. Оказалось, что афалины способны без специального обучения понимать логически упорядоченные последовательности "слов" языка-посредника. Интересно отметить, что постоянный оппонент Хермана Р. Шустерман (Schusterman, Gisiner, 1988) регулярно предлагал трактовку тех же результатов с позиций условнорефлекторной теории, отказываясь (подобно критикам экспериментов с "говорящими" обезьянами) признавать наличие у них хотя бы зачатков способности понимать значение порядка слов.
Несомненный интерес представляют также исследования на попугаях. Известно, что попугаи разных видов могут выучивать и произносить сотни слов, варьировать слова в предложениях, произносить фразы адекватно ситуации и вступать в довольно осмысленные диалоги со своими воспитателями. Подробное описание поведения "говорящих" птиц приводится в работе В.Д. Ильичева и О.Л. Силаевой (1990). Примеры "осмысленной" речи попугаев широко известны, особенно благодаря тому, что, начиная с 60-х годов, волнистые попугайчики стали широко распространенным у любителей видом. Однако экспериментальных исследований высокой нервной деятельности попугаев практически не проводится. Исключение составляют только многолетние работы американской исследовательницы Айрин Пепперберг (Pepperberg, 1981; 1987а; 1987Ь). На основе методик обучения обезьян она разработала оригинальный метод общения с попугаем Алексом (африканским серым жако — Psittacus erithacus), причем в качестве языка-посредника использовалась собственно человеческая речь. Хотя высказывания попугая в опытах Пепперберг и производят глубокое впечатление, их уровень не сравним с продемонстрированным шимпанзе. Тем не менее, именно Пепперберг впервые удалось поставить эксперименты, с помощью которых можно объективно судить о характере когнитивных способностей попугаев.
Разработанный Пепперберг метод отличается тем, что в процессе обучения участвуют одновременно два обучающих человека. Один (основной) тренер обращается одновременно и к человеку (второму тренеру), и к попугаю. Второй тренер является для попугая, с одной стороны, объектом для подражания, а с другой — как бы его соперником.
Благодаря этой программе Алекс научился не только называть предметы, но и определять их форму (треугольная, четырехугольная) и цвет, а также указывать материал, из которого они сделаны. Алекс мог отвечать на вопросы, например: "Сколько здесь предметов? Сколько из них круглых? А сколько кожаных? Сколько черных?" У этого попугая удалось установить связь между неприятной для него ситуацией и отрицанием "нет".
Работы по обучению языкам позвоночных-неприматов построены так, что не столько выявляют их коммуникативные способности, сколько характеризуют уровень когнитивной деятельности — способность к обобщению и символизации. Несмотря на то что такие работы немногочисленны и разрозненны, они внесли несомненный вклад в характеристику их высшей нервной деятельности и свидетельствуют о перспективности дальнейших исследований в этом направлении.
Чтобы точнее охарактеризовать структуру и свойства "языка", который усваивали обезьяны и другие позвоночные, кратко рассмотрим некоторые положения лингвистики, привлекаемые авторами экспериментов к их анализу, и определим их наличие или отсутствие в усваиваемых шимпанзе языковых навыках. Как упоминалось выше, существует много определений и критериев языка. В связи с проблемой усвоения антропоидами языков-посредников широко обсуждалось и использовалось определение известного американского лингвиста Ч. Хоккета (Hockett, 1958; см. также: Линден, 1981). Согласно его представлениям, язык человека обладает 7 ключевыми свойствами, которые частично присущи естественным языкам животных, а в полном объеме — только человеку. Использование предложенной Хоккетом базовой таблицы ключевых свойств позволяло понять, насколько глубоко животные могут овладеть человеческим языком и каковы пределы, их лингвистических способностей. В частности, Р. Футе (Fouts, 1975) проанализировал жестовый язык шимпанзе в соответствии с этими критериями.
Ч. Хоккет обсуждает выделенные им ключевые свойства языка применительно как к общению людей, так и к конкретным способам общения у четырех видов животных — пчелы, колюшки, серебристой чайки и гиббона, полагая, что такой подбор видов отражает разнообразие способов общения в животном мире. Как было описано выше, пчела, обнаружив источник пыльцы, возвращается в улей и танцем сообщает остальным пчелам о местоположении источника корма и о его количестве.
Самцы и самки колюшки сигнализируют о готовности приступить к размножению путем изменения своей окраски и формы тела. Птенцы серебристой чайки побуждают своих родителей кормить их с помощью клевков в направлении родительского клюва. В сообществе гиббонов существует система криков, оповещающих о различных опасностях и общих потребностях. Хоккет полагает, что в каждом из этих способов общения присутствует по крайней мере одно из выделенных им ключевых свойств языка, а в танце пчел — почти все (табл. 5.3, подробнее см.: Линден, 1981).
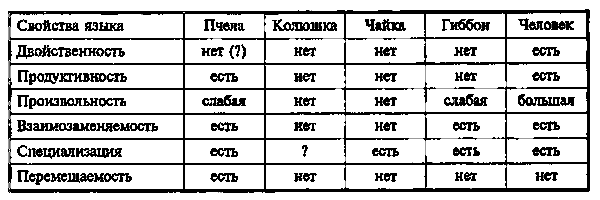
Таблица 5.3 Характеристики коммуникативных систем некоторых животных и человека.
Свойство структурной двойственности означает, что человеческий язык обладает одновременно и фонологической (звуковой), и грамматической (смысловой) организацией, т.е. построен из звуковых и смысловых элементов. Вместо того чтобы для каждого сообщения использовать отдельный сигнал, как это характерно для большинства коммуникативных систем животных, речь человека строится из конечного числа звуков, или фонем, которые комбинируются огромным числом различных способов и благодаря этому образуют смысловую структуру. Языки животных, как показано в таблице, этим свойством не обладают.
Свойство семантичностн означает присвоение определенного значения некоторому абстрактному символу. Это свойство присуще как естественным языкам животных, так и языкам-посредникам. Свойство произвольности означает, что сообщения слагаются из единиц, не имеющих очевидной связи с их смыслом. Альтернативой такой системе является иконическая, когда знаки являются "картинками", иллюстрирующими их смысл. Произвольность, как и двойственность, присуща и амслену, и йеркишу, и жетонам, с помощью которых "разговаривала" Сара (см.: 2.7.4). Примером иконического знака может служить изобретенный Уошо жест "нагрудник" (она очерчивала пальцем соответствующее место на груди), тогда как ее учили произвольному знаку — условной комбинации пальцев, ничего общего не имеющей с реальным предметом.
Свойство взаимозаменяемости заключается в том, что любой организм, способный посылать сообщения, должен быть способен и принимать их. Например, когда самка колюшки раздувает брюшко, самец в ответ совершает брачный ритуал, но их роли поменяться не могут, и таким образом, в языке колюшки взаимозаменяемость отсутствует. В отличие от этого все виды языков-посредников, освоенных шимпанзе, обладают этим свойством.
Свойство специализации заключается в отсутствии прямых физических следствий полученного сообщения, в том, что животное лишь передает информацию, но не действует непосредственно. Человек, по-видимому, обладает максимально специализированной системой общения. Если же вернуться к примеру с колюшкой, то самцы реагируют непосредственно на физические аспекты сообщения, посылаемого самкой, — раздувание брюшка и метание икры, тогда как самка реагирует на изменение окраски самца. Таким образом, сообщение самца более специализированно, чем сообщение самки. В отличие от естественных языков животных языки-посредники обладают свойством специализации.
Наконец, культурная преемственность означает способность передавать информацию о смысле сигналов из поколения в поколение посредством обучения и подражания, а не генетической преемственности. Наблюдения Р. Футса за общением Уошо с ее приемным сыном Лулисом (Fouts et al., 1984; 1989) показывают, что такая преемственность в использовании языка-посредника, по-видимому, может существовать.
Зарегистрировано, по крайней мере, три случая, когда Уошо специально учила малыша знакам амслена (пища, жвачка, стул) — складывала соответствующим образом его пальцы. Два этих жеста так и вошли в его словарь. Взрослые шимпанзе также в ряде случаев усваивали знаки, подражая "говорящим" сородичам.
Эти данные представляют несомненный интерес, однако не могут служить достаточно убедительным доказательством наличия культурной преемственности языковых навыков у шимпанзе. Хотя они пользуются знаками в отсутствие человека, неясно, насколько эти знаки отличаются по своим функциям от естественного языка жестов и телодвижений. Не было проанализировано, о чем обезьяны сигнализируют друг другу и какой тип коммуникативных связей они обеспечивают этими жестами. Вместе с тем, в природных условиях культурная преемственность, по-видимому, играет определенную роль в создании диалектов естественного языка шимпанзе.
Принципиально важным свойством языка человека является перемещаемость, поэтому мы особенно подробно рассмотрим, насколько оно присуще языковому поведению шимпанзе. Имеется в виду, что предмет сообщения и его результаты могут быть удалены во времени и пространстве от источника сообщения. Наличие этого свойства подразумевает способность:
1) использовать знаки в отсутствие соответствующего объекта;
2) передавать информацию о прошлых и будущих событиях;
3) передавать информацию, которая может стать известной только в результате употребления знаков.
У естественных коммуникативных систем животных свойство перемещаемости полностью отсутствует. В работах Р. Футса (Fouts, 1984) приведены отдельные наблюдения, свидетельствующие о наличии этого свойства в языке, усвоенном Уошо и Люси. Так, например, Люси, которую разлучили с ее любимой собакой, когда та заболела, постоянно называла ее по имени и говорила, что той больно (Temerlin, 1975).
Для анализа этого вопроса Р. Футc (Fouts et al., 1976) провел опыты на шимпанзе Элли. Он обратил внимание, что Элли неплохо понимает устную речь окружающих, и в специальном эксперименте обучил его названиям нескольких предметов. Затем Элли уже в отсутствие обозначаемых предметов научили знакам амслена, соответствующим этим словам. Во время теста обезьяне предъявляли новые предметы тех же категорий, что и использованные при обучении. Оказалось, что Элли правильно называл их с помощью жестов амслена, как бы мысленно переводя их названия с английского на амслен. На этом основании автор сделал заключение о том, что в основе употребления знаков у шимпанзе может лежать не просто образование ассоциаций, но формирование внутренних представлений о соответствующих им предметах и действиях. Поэтому знаки могут употребляться в отсутствие обозначаемых предметов и наряду с прочими преобразованиями допускают и кроссмодальный перенос от звуковых (словесных) к зрительным (жестовым) знакам.
Наиболее убедительно способность шимпанзе передавать информацию об отсутствующих и недоступных непосредственной сенсорной оценке предметах была продемонстрирована в работах С. Сэведж-Рамбо (Savage-Rumbaugh, 1984; 1993). При сравнении поведения шимпанзе и детей обращает на себя внимание тот факт, что дети понимают слова еще до того, как начинают говорить сами (навык восприятия формируется у них раньше, чем навык воспроизведения). В то же время этого не наблюдалось в опытах на многих шимпанзе, в особенности обученных "йеркишу" по методике Д. Рамбо. В этих опытах употребление знаков было жестко формализовано и служило главным образом для получения нужных предметов. В ряде случаев животные, по-видимому, не понимали смысла знаков при изменении контекста. Кроме того, в упомянутых выше опытах шимпанзе подают знаки главным образом в присутствии предметов, если они получают за это подкрепление, тогда как дети произносят слова ради чистого удовольствия или для привлечения внимания окружающих к интересующим их вещам.
С. Сэведж-Рамбо предположила, что эти различия обусловлены особенностями методики, которую разработал для обучения шимпанзе Д. Рамбо. По ее мнению, ведущую роль в ней играло образование условной связи (ассоциации) между знаком и получением соответствующего предмета, а также подражание действиям тренеров, тогда как у детей слова усваиваются в самых разных ситуациях и им соответствуют более или менее отвлеченные внутренние представления, а не простые ассоциации. Особенно это различие проявлялось при сравнении поведения детей и шимпанзе Даны. Ее обучение было строго формализовано во избежание возможных подсказок и т.п., и она действительно научилась главным образом нажатию на соответствующую клавишу компьютера для получения нужного ей предмета.
В отличие от этого новая методика С. Сэведж-Рамбо не фокусировала внимания обезьян на получении объекта. Их учили использовать знак (изображение, которое появлялось на экране после нажатия на определенную клавишу, так называемая "лексиграмма") не для получения предмета, а для его "наименования" в ситуации, где всегда было несколько предметов, и воспроизведение знака не было жестко связано с их получением. Поскольку одновременно воспитывали двух самцов — Шермана и Остина, — они научились общаться с помощью знаков, отвечать на них и влиять на поведение друг друга и окружающих. Благодаря этому они, по мнению авторов, усваивали язык как "функционально-действенный навык", как средство воздействия на поведение окружающих, а не просто как серию движений, которую надо совершить, когда этого хочет экспериментатор. С нашей точки зрения, сходный тип поведения был описан Футсом (Fouts et al., 1984) у шимпанзе Бруно и Буи, которых обучали амслену (см. табл. 5.2).
Обученные по этой системе Шерман и Остин употребляли знаки в гораздо более разнообразных ситуациях, чем их предшественники, особенно Дана. Стремление к "наименованию" предметов обнаружилось у них спонтанно, без инструкции тренера. Очевидно, что научить этому специально невозможно. Не дожидаясь, когда тренер даст им или попросит предмет, как это бывало у всех других обезьян, они по собственной инициативе называли их и показывали тренеру, т.е. включали обычно исполняемые тренером функции в собственное поведение. При появлении тренера с набором игрушек они без всякой команды называли их и показывали, вместо того, чтобы ждать, когда тот решит играть и даст соответствующую команду. Это свойство проявилось и в отношениях между самими шимпанзе. Шерман — доминант — нажимал на соответствующую клавишу и давал Остину кусок апельсина. Оба нажимали нужную клавишу компьютера прежде, чем взять кусок пищи или игрушку, даже если те были спрятаны. Остин (низший по рангу) подводил Шермана к компьютеру и побуждал добывать лакомства.
Как подчеркивает автор, для такого уровня использования знаков (лексиграмм) как средства индикации (называния) предметов требуется понимание соответствия между: 1) предметом, который выбран из группы как объект наименования; 2) лексиграммой, выбранной на панели; 3) предметом, который в конце концов выбран и передается экспериментатору. Наиболее жесткая проверка этого соответствия — процедура теста при двойном слепом контроле (рис. 5.24).
Рис. 5.24. Тест на символизацию при двойном слепом контроле. 1 — стол с предметом; 2 — клавиатура для выбора; 3 — экспериментатор, оценивающий результат выбора (пояснения в тексте)
В обстановке игры с тренером шимпанзе "называют" один из предметов, но предметы для выбора, тренер, которому их надо показывать, и клавиатура, на которую надо нажимать, пространственно разнесены для создания дополнительной нагрузки на образную память. Решив, какую игрушку выбрать (рис. 5.24, а), шимпанзе должен помнить, что именно он выбрал, пока идет к клавиатуре и высвечивает соответствующую лексиграмму в отсутствие реального объекта. Затем он должен держать в памяти высвеченную лексиграмму, когда перестает ее видеть, взять соответствующий предмет (рис. 5.24, б) и передать его тренеру, который сидит у экрана и видит лексиграмму только после вручения ему выбранного предмета, так что не может ничего "подсказать" обезьяне (рис. 5.24, в).
Оказалось, что оба шимпанзе правильно называли выбранный предмет практически в 100%, причем, когда Шерман однажды высветил лексиграмму отсутствующего в наборе предмета, то не взял ничего. Эти опыты позволили сделать важное заключение о языковых возможностях шимпанзе.
По мнению авторов, стремление Шермана и Остина к наименованию предметов по собственной инициативе, в том числе когда они их не видят, свидетельствует о том, что усвоенные ими знаки приобрели свойства символов и эквивалентны навыкам, описанным у детей. Они несомненно свидетельствуют о том, что языковые навыки обезьян при определенных условиях обучения могут обладать свойством перемещаемости. Еще более веские доказательства были получены в дальнейших работах С. Сэведж-Рамбо на карликовом шимпанзе Кэнзи (см.: 5.4.4.4).
Как мы уже упоминали, вопрос о наличии свойства перемещаемости в усвоенном шимпанзе языке особенно важен в связи с изучением мышления животных, поскольку употребление знака в отсутствие обозначаемого им предмета свидетельствует о формировании и хранении в мозге внутренних (мысленных) представлений об этом предмете.
Свойство продуктивности означает способность создавать и понимать бесконечное число сообщений, преобразуя исходный ограниченный запас символов в новые сообщения. Именно это свойство делает языки человека практически не ограниченными по своему объему и богатству. Вопрос о наличии этого свойства у языковых навыков шимпанзе был многократно проанализирован. О его наличии могла свидетельствовать способность комбинировать знаки для обозначения новых предметов: например, Уошо называла арбуз "candy drink' ("конфета — питье"), а впервые встреченного на прогулке лебедя — "water bird" ("вода — птица"). При достаточно большом запасе знаков шимпанзе начинали гибко использовать синонимы для обозначения одного и того же предмета в зависимости от контекста (чашка — пить, красный, стекло; подробнее см.: Линден, 1981. С. 104-110).
Приведенные данные, по-видимому, свидетельствуют, что усвоенная шимпанзе система знаков амслена в некоторой степени обладает свойством "продуктивности". Впоследствии анализу этого свойства языков-посредников специального внимания почти не уделялось. Что касается естественных языков животных, то до недавнего времени считалось, что у них это свойство практически отсутствует. В большинстве случаев это действительно так, однако ряд современных исследований показывает, что один компонент естественного языка шимпанзе — так называемые "долгие крики" — состоит из ограниченного числа базовых элементов, которые комбинируются по-разному в зависимости от ситуации и у разных животных и тем самым расширяют диапазон возможных сообщений (Mittaiii, 1992; Clark, Wrangham, 1993; Brandt, 1994; Hofman, Fruth, 1994).
В.И. Марков (1993) на основе многолетнего анализа акустических сигналов дельфинов приходит к выводу, что они обладают коммуникативной системой открытого типа, которая также наделена свойством продуктивности, хотя вопрос этот, несомненно, требует дальнейших исследований. Таким образом, в отличие от большинства естественных коммуникативных систем животных языки-посредники, которыми овладевали шимпанзе, обладают не только свойством семантичности, но и элементами свойств продуктивности и перемещаемости.
Уже на самых ранних этапах экспериментов выяснилось, что обезьяны комбинировали знаки не только для обозначения новых предметов. Оказалось, что они склонны спонтанно объединять 2—3 и более знаков в цепочки, напоминавшие предложения, которые произносят дети 1,5—2,5 лет. Огромная доля усилий Гарднеров, их коллег и последователей была направлена на анализ структуры создаваемых обезьянами "предложений". Анализ 158 фраз, "произнесенных" Уошо, показал, что в большинстве случаев порядок слов в них отвечает принятому в английском языке и отражает те же, что у детей, базисные семантические отношения типа: субъект — действие, действие — объект, указательная частица — объект. Иными словами, обезьяны прекрасно понимали и передавали информацию о направленности действия, о принадлежности предмета, о его местонахождении ("Роджер щекотать Люси", "дай мне, я дам тебе", "кошка кусать собака" или "собака кусать кошка" и т.п.). Таким образом, типичные для шимпанзе последовательности знаков обычно были основаны на улавливании связей между предметами и явлениями внешнего мира, отражали их эмпирические представления.
На основании этих данных было высказано предположение о том, что обезьяны овладевают элементами синтаксиса. Более того, ряд авторов истолковывает эти данные как подтверждение гипотезы американского лингвиста Ч. Хомски (Chomsky, 1980) о наличии неких врожденных "языковых структур", т.е. механизмов, которые предопределяют освоение языка человеком в процессе его онтогенеза.
Эта гипотеза Хомски подвергалась широкой критике (см.: Панов, 1983), но в определенный период ее использование было продуктивным при анализе поведения обезьян в процессе освоения языка.
Вопрос о синтаксисе языка Уошо и других шимпанзе долгое время был предметом острых дискуссий. Позднейший анализ тех же видео- и кинозаписей показал, что первоначальные представления были слишком оптимистичными и не полностью подтверждались. Так, оказалось, что увеличение числа знаков в предложении чаще всего не прибавляет объема передаваемой обезьяной информации, что многие из фраз остаются незаконченными, а часть из них — бессмысленными. Однако выяснилось, что в лингвистике и детской психологии не существовало критериев того, с какого момента, с какой стадии детский лепет можно считать речью. Это и понятно, поскольку не было такой проблемы: ведь у детей все равно раньше или позже этот лепет переходил в полноценную речь. Но для строгого сравнительного анализа речи ребенка и шимпанзе такие критерии были необходимы.
Одним из наиболее непримиримых противников гипотезы о том, что "язык" обезьян обладает синтаксисом, был и остается до настоящего времени американский исследователь Герберт Террес (Terrace et al., 1979). Он обучил шимпанзе Нима амслену по методике Гарднеров и проанализировал его высказывания, сравнив их с фразами Уошо. По убеждению Терреса, данный уровень экспериментов не позволяет утверждать, что шимпанзе действительно овладевают элементами синтаксиса, т.е. понимают смысл знаков и употребляют их в определенном порядке в соответствии с законами английской грамматики. По его мнению, в основе создаваемых обезьянами фраз лежит не истинное представление о грамматических структурах, а какой-то более простой и, вероятно, более универсальный механизм.
Таким механизмом, как считал Г. Террес, может быть, например, способность животных подражать экспериментатору: они запоминают продуцируемые им последовательности знаков и воспроизводят их в соответствующей ситуации. Он предположил, что запоминание длинных цепей знаков — это особый процесс, отличный от механического заучивания. В его основе, по-видимому, лежит улавливание их внутренней структуры (Terrace 1985; 1991). Этот процесс можно сравнить с запоминанием, например, телефонных номеров, когда цифры группируют парами или тройками.
Одно из наиболее весомых возражений Терреса состояло в том, что, по его мнению, обезьяны строили свои фразы просто в подражание воспитателям. Поскольку Гарднеры не регистрировали жестов экспериментаторов, они не могли удовлетворительно опровергнуть этот довод (или согласиться с ним). Они повторили работу с 4 новыми обезьянами (Gardner et al., 1989), однако не сделали попытки определить в ответ на критику, влияют ли адресованные обезьянам фразы тренеров на их жестикуляцию. В новой работе они еще раз показали, что и эти новые 4 шимпанзе прошли тесты двойного слепого контроля, подобные тем, которые проводили с Уошо. Эти тесты ясно свидетельствуют, что шимпанзе могут узнавать предметы и действия на фотографии, продуцируя соответствующие знаки в условиях, исключающих подсказку.
Однако Терресс никогда и не оспаривал весомости этих тестов: его критические замечания были направлены на то, что обезьяны вряд ли способны самостоятельно формировать семантически значимые и грамматические правильные предложения, не подражая своим тренерам. К сожалению, Гарднеры так и не дали убедительного ответа на этот вопрос. Веские доказательства того, что шимпанзе действительно могут усваивать общие принципы построения фраз и даже делать это самостоятельно, на основе понимания их смысла, а не просто подражая экспериментатору, были получены лишь в работах Сью Сэведж-Рамбо (Savage-Rumbaugh S.E. et al., 1993) в 90-е годы.
Выше упоминались работы (см.: 5.4.4.3), в которых шимпанзе Шерман и Остин обнаружили способность употреблять символы для обозначения отсутствующих предметов и в более широком контексте, чем обученные по другим методикам. В развитие этой работы Сэведж-Рамбо начала воспитывать детенышей карликового шимпанзе и детей в сходной речевой среде и сравнила их способности понимать устную речь. Особенность этого долгосрочного эксперимента состояла в том, что он был начат в раннем возрасте, когда двум карликовым шимпанзе (бонобо) и трем детям было по 10 месяцев. Все они с начала обучения находились в сходных условиях: постоянно слышали устную человеческую речь. Шимпанзе научились также общаться с человеком и друг с другом при помощи компьютера. При этом обезьян не тренировали непосредственно на выполнение словесных команд, а лишь создавали им "языковую среду".
Оказалось, что в возрасте 5 лет Канзи спонтанно начал понимать устную речь, и не только отдельные слова (что отмечали и раньше другие исследователи), но и целые фразы. Без специальной тренировки он правильно выполнял каждый раз новые команды и понимал разницу между фразами "Унеси картошку за дверь" и "Иди за дверь, принеси, картошку".
Опыты убедительно свидетельствуют, что шимпанзе могут понимать и сложные синтаксические конструкции типа: "Если не хочешь яблока, положи его обратно". Подобные конструкции понимала и строила также шимпанзе Сара в опытах Д. Примэка (A. Premack, D. Premack, 1971, 1994). Однако она делала это только после долгой тренировки с каждой конкретной фразой, не понимая общего принципа, тогда как Кэнзи усваивал именно общий принцип и без дальнейшей дрессировки спонтанно мог давать собственные ответы на сотни вопросов. Кэнзи научился также общаться с человеком и другими обезьянами при помощи лексиграмм, наблюдая, как этому учили его приемную мать Матату (причем безуспешно). Свои собственные фразы он также составлял в соответствии с правилами, так что, например, лексиграмма, обозначающая его приемную мать, оказывалась то подлежащим, то дополнением ("Хватай Матату", "Матата кусает"). Вывод о том, что шимпанзе действительно улавливает синтаксический строй английской речи, был сделан на основе экзамена, который включал 660 вопросов, заданных устно и совершенно не повторяющихся. Оказалось, что, как и его двухлетняя коллега — девочка Аля, он безошибочно понимал все предложенные ему просьбы и вопросы (рис. 5.25).

Рис. 5-25. Понимание устной речи у шимпанзе (рисунок Н. Рябчиковой). По команде "Сделай собаке укол" Кэнзи приносит плюшевого щенка, потом находит шприц, снимает с него колпачок и колет им собаку, В другой раз в ответ на просьбу "Пусть собака укусит змею" он находит среди прочих предметов игрушечную змею, открывает рот собаке и изображает укус
На основе этих опытов Сэведж-Рамбо сделала вывод, что язык Кэнзи достигает "протограмматического уровня, сравнимого по сложности с речевыми навыками двухлетнего ребенка". Еще один важный вывод работы состоит в том, что и для человека, и для шимпанзе критическим фактором, определяющим формирование способности понимать речь, является возраст, когда они начинают слышать речь, и условия, в которых они при этом находятся. Ведь шимпанзе, которых начали обучать не в 10 месяцев (как Кэнзи), а в 2—3 года, смогли усвоить гораздо более ограниченные навыки и не были способны понимать устную речь.
Эти сенсационные данные заставили авторов обратиться к исследованию мозга шимпанзе в поисках морфологических основ зачатков речи. С помощью разнообразных методов (гистология, сканирование, позитронно-эмиссионная томография) была обнаружена асимметрия височных структур, причем в левом полушарии область planum temporale оказалась более развитой, чем в правом. Выводы американских ученых представляются тем более убедительными, что еще в 30—60-е годы российские нейроморфологи Г. И. Поляков, С.М. Блинков, Е.П. Кононова, Ю.Н. Шевченко (см.: Фирсов, 1993) разрушили представление о том, что речевые зоны специфичны только для мозга человека. Они обнаружили многочисленные черты сходства речевых областей мозга человека и шимпанзе. В частности, еще в тот период было установлено, что в верхневисочной области шимпанзе имеется поле 37, ответственное у человека за понимание звуковой речи. Коль скоро мозг шимпанзе наделен речевыми структурами и способен их активизировать в соответствующих условиях, можно предположить, что последний общий предок человека и шимпанзе тоже имел эти структуры. Тогда и непосредственные предшественники человека — австралопитеки и Homo erectus — тоже могли иметь зачатки языка (Savage-Rumbaugh et al., 1993).
Сколь ни убедительны были бы данные о способности шимпанзе к усвоению и пониманию синтаксиса языков-посредников, их значение несомненно возрастет, если в естественном поведении шимпанзе удастся обнаружить черты, соответствующие полученным в лаборатории. Для этого необходимо выяснить, например, используются ли в естественном языке шимпанзе специфические ряды звуковых элементов, которые меняют свой смысл в зависимости от того, в каком порядке эти элементы следуют друг за другом. Такой факт свидетельствовал бы, что они организованы в соответствии с неким "естественным синтаксисом". Исследования этого аспекта коммуникации приматов оказались достаточно плодотворными. Показано, что разная комбинация одних и тех же элементов обеспечивает различия между криками самцов и самок у тамаринов. У гиббонов репертуар богаче и может быть дифференцирован более тонко. Так называемые "долгие крики" сигнализируют не только о присутствии обезьяны и ее местонахождении, но также о ее поле, ранге и "семейном положении" (Cowlishaw, 1992).
В репертуаре обоих видов шимпанзе (и обыкновенного, и карликового, или бонобо) также существуют "долгие крики". Они состоят из ограниченного числа базовых акустических элементов, которые комбинируются по-разному у разных животных (Clark, Wrangham, 1993; Mitani, Brandt, 1994; Hofman, Fruth, 1994).
Важную роль в общении приматов играют "дуэты" — координированные крики, когда обезьяны взаимодействуют, подстраиваясь под голоса друг друга. У гиббонов дуэты устойчивы и однообразны. Половые различия, похоже, генетически запрограммированы. В криках бонобо есть различия, но оба пола способны продуцировать звуки одной частоты, и когда они кричат по одиночке, их трудно различить (Hohmarm, Fruth, 1994).
Митани и Брандт (Mitani, Brandt, 1994) наблюдали, что самцы шимпанзе пытаются подражать акустическим характеристикам криков обезьяны, с которой они кричат вместе. Отдельные самцы, как оказалось, кричат по-разному в зависимости от того, с какими партнерами взаимодействуют. Это приводит, с одной стороны, к индивидуальному разнообразию, но с другой — к унификации криков членов данной группы.
Индивидуальный репертуар каждого самца содержит много вариантов криков, которые усваиваются благодаря подражанию сородичам. Как считают авторы, это свойство отличает язык шимпанзе от обычных коммуникативных систем животных. На основе этих данных высказано предположение, что естественная коммуникативная система шимпанзе может рассматриваться как промежуточная между обычными для животных и языком человека. Для ее обозначения введен термин "минимальный язык" (или "протоязык"), имеющий в виду наличие в криках шимпанзе вариабельных элементов, которые по-разному группируются в разных ситуациях (элементарный синтаксис), и не только выражают сиюминутное состояние животного, но сигнализируют о самоопределении особи.
Сходное мнение о том, что у шимпанзе может иметься некий "первичный язык", высказывает Л.А. Фирсов (1993) на основе всестороннего изучения высшей нервной деятельности шимпанзе. Предполагается, что для возникновения этих особенностей коммуникации существенна сложная сеть социальных взаимодействии, характерная для сообществ шимпанзе, поскольку ни моногамные виды, ни виды, живущие в сообществах другого типа, не могут достичь подобного уровня.
Таким образом, есть данные о том, что естественный язык шимпанзе качественно отличен от коммуникативных систем других приматов. Эти данные также подтверждают уже упоминавшиеся нами представления Л.А. Орбели (1949) и О. Келера (Koehler, 1956) о наличии промежуточных, переходных уровней коммуникации, предшествовавших появлению речи человека. В сочетании с результатами обучения приматов языкам-посредникам, сконструированным на базе человеческой речи, на этом основании можно сделать вывод о столь значительных резервах коммуникативных, а также когнитивных возможностей животных, какие трудно было представить себе до начала 70-х годов.
В завершение обзора проблемы языков животных рассмотрим еще один, принципиально иной подход к исследованию языка животных, основанный на идеях теории информации (Резникова, 1997; Резникова, Рябко, 1988, 1990, 1995а, б). Суть его в том, что в экспериментах испытуемым предлагалось передать заранее известное экспериментатору количество информации. При этом измерялось затраченное испытуемым время, т.е. оценивалась скорость передачи информации. Авторы исходят из того, что кроме перечисленных выше свойств языка по Хоккету, таких как продуктивность, семантичность, перемещаемость, язык должен обладать еще одним свойством: размер сообщения должен быть пропорционален количеству информации в нем. Поясним это требование. После введенного К. Шенноном в конце 40-х годов строгого понятия "количество информации" были исследованы многие естественные языки человека, и во всех них длина сообщения оказалась пропорциональной количеству содержащейся в нем информации. Иными словами, на двух страницах книги можно разместить в два раза больше сведений, чем на одной.
Что же такое информация по Шеннону? В опыте "орел или решка" возможны два равновероятных исхода: подброшенная монета падает вверх либо гербом, либо цифрой. Если кто-нибудь сообщит нам результат такого опыта, он передаст 1 бит информации (бит — единица измерения информации). Вообще, если опыт имеет n равновероятных исходов, и нам сообщают его результат, то мы получаем log 2n битов информации. На рассмотренном понятии информации основаны современная теория и практика построения систем связи (основополагающая работа Шеннона так и называлась — "Математическая теория связи"). В дальнейшем выяснилось, что это же понятие играет фундаментальную роль в психологии, лингвистике и других областях. Благодаря этому систему коммуникации животных удалось проанализировать как средство передачи информации — конкретной, количественно измеримой величины. Объектом исследования служили муравьи. В 1899 г. немецкий зоолог Е. Васман предложил гипотезу так называемого антеннального кода — своеобразного языка жестов, основанного на тонких движениях антенн (усиков). О том, как выглядит предполагаемый процесс обмена информацией у многих видов муравьев, образно написал Данте в "Божественной комедии":
".. .Так муравьи, столкнувшись где-нибудь,
Потрутся рыльцами, чтобы дознаться,
Быть может, про добычу иль про путь...
Но только миг объятья дружбы длятся."
Несмотря на большое число работ, в том числе основанных на методах макросъемки, существование у муравьев языка, подобного, например, языку танца у медоносных пчел, или вообще системы дистанционного наведения на добычу, оставалось недоказанным.
В описываемых ниже опытах муравьи могли получить пищу лишь в том случае, если они передавали друг другу заданное экспериментатором количество информации. Для этого муравьев помещали в специальный лабиринт, смонтированный из спичек и названный "бинарным деревом". В простейшем случае он имел одну развилку, в вершинах которой находились кормушки: одна пустая, другая — с сиропом. Чтобы другие муравьи могли найти корм, разведчики должны были сообщить им "иди налево" или "иди направо", т.е. передать 1 бит информации. Максимальное число развилок лабиринта в некоторых опытах доходило до 6, и только в вершине одной из них находилась кормушка с сиропом, остальные были пустыми. В таких опытах муравьи могли быстро отыскать корм, если получали сведения о последовательности поворотов типа "ЛПЛППЛ" (налево, направо и т.п.).
При 6 развилках в лабиринте им необходимо было передать 6 битов информации. Муравьи разных видов содержались в прозрачных гнездах из оргстекла на больших лабораторных аренах, и можно было наблюдать любые контакты между ними. Чтобы знать муравьев "в лицо", их метили индивидуальными цветными метками.
Оказалось, что при решении сложных задач среди муравьев выделяются постоянные по составу рабочие группы, состоящие из одного разведчика и 4—7 фуражиров. Разведчик, найдя пищу, подзывает к ней только свою группу фуражиров. В эксперименте муравьи получали пищу раз в 3 дня и только в лабиринте, т.е. их поиски стимулировались голодом. Лабиринт укрепляли в кювете с водой, чтобы муравьи не могли двигаться по прямой, а в исходный пункт лабиринта попадали бы с арены по мостику. Муравьи, находящиеся в гнезде и на арене, не могли наблюдать за действиями разведчика в лабиринте. Чтобы исключить возможность использования пахучего следа, лабиринт заменяли идентичным в то время, когда разведчик находился в гнезде. Более того, все кормушки в нем были только пустыми (в случае правильного выбора муравьев срочно кормили), так что исключалась и ориентация муравьев по запаху сиропа. В каждом сеансе измерялась общая длительность контактов фуражиров с разведчиками, вернувшимися в гнездо после обнаружения пищи. При этом фуражиры были вынуждены действовать самостоятельно: разведчика изымали пинцетом и временно отсаживали в баночку.
Выяснилось, что по способности запоминать и передавать информацию разведчики различаются. "Бестолковые" разведчики не могут сообщить правильную информацию, поэтому направляемые ими группы фуражиров идут по неверному пути и, как правило, возвращаются в гнездо, не достигнув цели. Естественно, таких разведчиков из эксперимента исключали, а использовали в нескольких сеансах "перспективных" разведчиков и группы работающих с ними фуражиров. Такие разведчики не только сразу запоминали дорогу, но и сравнительно легко переучивались, когда в ходе сеанса менялось направление пути к приманке, т.е. если ее переставляли на вершину другой ветки того же бинарного дерева. Разведчиков, отобранных в первых, более простых опытах, в дальнейшем просто сажали на вершины с приманкой. Прежде чем мобилизовать свою группу, разведчик совершал один—два, а иногда и больше рейсов от приманки до гнезда, запоминая путь. Наблюдая за его возбужденными действиями и получая от него пищу, группа постепенно подтягивалась к выходу.
В экспериментах фиксировали длительность контакта разведчика и фуражиров с помощью секундомера. Началом антеннального контакта разведчика с фуражирами считалось его прикосновение к первому муравью, а окончанием — выход из гнезда первых трех фуражиров. После такого контакта фуражиры компактной группой безошибочно направлялись к приманке и находили ее менее чем за минуту. Случалось, что тот или иной муравей уклонялся на ложный путь, но довольно быстро возвращался к своей группе. В контрольных опытах фуражиры, а также опытные разведчики, допущенные в лабиринт, но не знакомые с координатами кормушки, не находили ее за отведенные им 15 мин. Это позволяет исключить действие каких-либо иных способов ориентации, кроме использования сведений, сообщенных разведчиком.
В опытах с бинарным деревом количество информации (в битах), необходимое для выбора правильного пути в лабиринте, равно числу развилок. Оказалось, что у трех видов муравьев с групповой организацией доставки пищи зависимость между временем контакта разведчика с фуражирами и количеством передаваемой информации (числом развилок) близка к линейной и описывается уравнением t = ai + b, где t — время контакта, а — коэффициент пропорциональности, равный скорости передачи информации (число битов в минуту). Постоянная b была введена нами потому, что муравьи могут передавать информацию, не имеющую прямого отношения к поставленной задаче, например, просто сигнализировать: "есть пища". Отметим сразу, что скорость передачи информации у муравьев, по крайней мере, на порядок ниже, чем у человека: около 1 бит в минуту.
Одной из важнейших характеристик языка и интеллекта его носителей следует считать способность быстро подмечать закономерности и использовать их для кодирования, т.е. "сжатия" информации. В таких случаях оказывается, что размер сообщения о некотором объекте или явлении должен быть тем меньше, чем они проще, т.е. чем больше в них закономерностей. Известно, например, что человеку легче запомнить и передать информацию о последовательности поворотов на пути к цели "ЛП ЛП ЛП ЛП ЛП ЛП ЛП" (налево-направо, и так 7 раз), чем о более короткой, но неупорядоченной последовательности "ПЛЛПППЛП". Оказалось, что язык муравьев и их интеллект позволяют им использовать простые закономерности "текста" для его сжатия (здесь "текст" — последовательность поворотов на пути к кормушке).
Примечательно, что муравьи начинают "сжимать" информацию только при достаточно больших "текстах", т.е. начиная с 4—5 развилок. Время, затраченное разведчиками на передачу информации о "закономерных" последовательностях поворотов, было значительно меньше затраченного на передачу "случайного" текста. Например, чтобы передать последовательность "ЛЛЛЛЛЛ", им требовалось вдвое меньше времени, чем для передачи последовательности "ПЛПППЛ". В общем случае время передачи сведений росло по мере усложнения "текста", т.е. наличия случайности, неупорядоченности в последовательностях поворотов на пути к кормушке.
Итак, выяснилось, что муравьи способны передавать друг другу довольно много различных сообщений, а время передачи сообщения пропорционально количеству информации в нем. Более того, они способны подмечать закономерности в передаваемой информации и использовать их для "сжатия" сообщения.
Последующие опыты были построены по тому же принципу, но с несколько измененной методикой. Экспериментальная установка напоминала длинную гребенку, размещенную в разных вариантах вертикально, горизонтально или замкнутой в круг, и муравьи должны были передавать сведения о номере планочки с приманкой. Эти опыты выявили у муравьев даже некую систему счисления, сходную с архаичными системами счисления у человека, когда информация о числе передавалась по типу "палец, палец, палец..." (подробно см.: Рябко, Резникова, 1997). Видимо, такая коммуникативная система по своим свойствам превосходит языки большинства животных. Заметим только, что среди огромного числа видов муравьев (их около 6 тысяч) подавляющее большинство не пользуется передачей столь сложной информации. У многих видов в естественных условиях семью снабжают одиночные фуражиры: небольшая часть семьи ведет активный поиск добычи на кормовом участке, справляясь со всеми задачами в одиночку или взаимодействуя через подражание. Другая, довольно большая группа видов использует другой способ. Немногочисленные разведчики мобилизуют на поиск пищи массу пассивных фуражиров с помощью запаховой сигнализации пахучего следа.
Для обеих групп видов размер кормового участка ограничен дальностью рейсов или невозможностью проложить пахучий след на большом расстоянии. Эти факторы накладывают ограничения на численность семьи: как правило, у таких видов она насчитывает от сотен до нескольких тысяч особей. В описываемых опытах представители упомянутых групп видов не использовали столь сложный способ обмена информацией.
Лишь немногие виды муравьев достигли высшего уровня социальной организации (описанного как "колонии" и "федерации", см.: 5.6.3) и максимально возможного для этой группы, когда они практически не имеют врагов, широко распространены и обладают огромной численностью и биомассой.
Только у представителей этих видов обнаружены описанные в данном разделе наиболее сложные формы коммуникации (Резникова, 1997). В заключение этого раздела отметим, что теоретико-информационный подход к исследованию языка животных можно использовать для анализа коммуникации не только муравьев, но и других общественных животных — термитов, дельфинов, обезьян. При этом, разумеется, техника экспериментов должна быть изменена с учетом особенностей поведения и размеров объектов исследования.
Свойственный каждому виду характер общественного поведения можно рассматривать как некий условный эталон, поскольку целый ряд факторов может вызвать значительные отклонения от него. Ввиду этого исследователи не оставляют попыток создать новые, более совершенные способы классификации сообществ по сравнению с рассмотренной выше общей схемой (подразделение сообществ по признаку "знание" или "незнание" особями друг друга).
В разделе 5.2 упоминалось о признаках, которые важны для классификации типов сообществ. Эту общую типологическую схему социальных систем разработал Е.Н. Панов (1983). Предлагаемая им классификация социальных систем у животных учитывает индивидуальное распознавание, выраженность территориального поведения, устойчивость группировок и т.д. Экологические факторы, влияющие на организацию сообществ, лучше всего исследованы у млекопитающих, однако выделить те из них, которые определяют величину группы и характер внутригрупповых взаимоотношений, оказалось далеко не просто. На первый план выступают два фактора: распределение пищевых ресурсов и давление со стороны хищников. Ниже приводятся краткие сведения об организации сообществ у некоторых видов, причем предпочтение отдается тем из них, которые наиболее подробно рассматриваются в связи с проблемами обучения, рассудочной деятельности или генетики поведения. Изложение ограничивается представителями отрядов грызунов, хищных приматов (позвоночные) и общественными насекомыми (беспозвоночные).
Группировки грызунов, достаточно разнообразные по своей структуре, могут в то же время служить примером ее пластичности. У грызунов разных видов структура сообщества тесно связана с экологической специализацией данного вида и/или популяции (см.: Шилов, 1977).
В целом можно сказать, что грызуны дают примеры всевозможных вариантов социальной организации от строгой обязательной территориальности (как у некоторых видов полевок) до больших колоний с более мелкими индивидуализированными группами и сложной системой акустической сигнализации (луговые собачки, сурки, пищухи). У голых землекопов (Heterocephalus glaber, подробнее см.: ниже) сообщество устроено по типу, сходному с эусоциальными сообществами общественных насекомых.
Серые крысы. Исследования иерархической организации и структуры сообществ серых крыс были предприняты еще в 50—60-е годы ввиду большой практической важности информации о биологии этого вида. Классическая работа Дж. Колхауна (Calhoun, 1962), проводившаяся в течение нескольких лет, содержала протоколы наблюдений за большой колонией крыс-пасюков, поселенных в очень большую вольеру.
Наблюдения показали, что колония пасюков — анонимное сообщество закрытого типа. Для нее характерно жесткое доминирование наиболее сильных самцов, вокруг которых группировались самки. Другие, более низкоранговые животные занимали менее удобные места, а самки, которые держались около них, приносили значительно меньшее количество детенышей.
Среда обитания серой крысы резко отличается от экологических условий, в которых живут другие виды грызунов. Этот факт необходимо учитывать при сопоставлении данных о социальном поведении серой крысы и грызунов других видов. Важно также отметить, что лабораторная белая крыса — альбиносная форма Rattus norvegicus — является наиболее популярным объектом при исследовании физиологии поведения (см. рис. 1.1). Работа Колхауна показала, что дикий родич лабораторной крысы обладает сложной системой врожденных реакций, позволяющей этим животным существовать в больших, пространственно разобщенных группах.
Знание биологии поведения этого вида в естественных условиях должно непременно учитываться при изучении поведения крыс в лаборатории. В работе Колхауна было показано, что даже при избытке пищевых ресурсов возрастание плотности популяции сопровождается снижением рождаемости.
Этот феномен обеспечивается целым рядом нейрогуморальных механизмов. Воздействие плотности популяции на состояние репродуктивной сферы у мышевидных грызунов было в дальнейшем изучено в работах Кристиана и Дэвиса, проведенных в 60—70-е годы, в дальнейшем эти данные многократно подтверждались другими исследователями.
Домовые мыши. Общественное поведение этих животных было блестяще описано в книге П. Кроукрофта "Артур, Билл и другие, ...или Все о мышах" (1972). Отличительной чертой сообщества домовой мыши является формирование большой семейной группировки, или "дема", члены которой держатся изолированно от других, ей подобных, и в пределах которой обычно производят потомство один или несколько доминантных самцов, а молодые самцы вытесняются и мигрируют в другие места. Установление иерархических отношений в искусственных группах самцов, исследования обонятельных сигналов, которые служат ключевыми раздражителями для запуска агрессивных реакций доминанта, и многие другие вопросы, связанные с видоспецифическим поведением домовой мыши, были исследованы на лабораторных инбредных линиях (Новиков, 1988; Maxson, 1992) и частично будут рассмотрены в главе 8.
Песчанки. Общественное поведение полуденной (Meriones meridianus), монгольской (М. unguiculatus), краснохвостой (M.libycus) и др. песчанок было исследовано в работах М.Е. Гольцмана с соавт. (1994).
Особенности организации сообществ у этих животных были проанализированы на нескольких уровнях: в полевых условиях, когда проводили наблюдение за жизнью колоний диких зверьков, в "макронаблюдениях" за искусственной популяцией животных в лаборатории, когда можно было детально проследить за динамикой иерархических отношений в семейных группах, и в детальных "микронаблюдениях" за особенностями микрокинематики ритуальных видоспецифических ФКД.
В естественных условиях жизни для монгольских песчанок характерен достаточно высокий уровень "общительности" и стремления вступать в контакты. Группировки особей (семейные группы) y этого вида сохраняются даже после переселения их на соседние участки. Социальные контакты особей этого вида весьма сложны и многообразны и характеризуется наличием четких поведенческих ролей (см. также: 5.3.7). Они отличаются абсолютным характером доминирования, а в лабораторных условиях — рождением детенышей только одной самкой при наличии нескольких самок в группе. В эксперименте (в условиях вольерного содержания) их характеризуют четкость и разнообразие поз и движений при агрессивных контактах, что сочетается с высокой стереотипностью поведения при взаимодействиях особей.
Краснохвостые песчанки в природных условиях избегают контактов друг с другом; контакты эти короче по времени и примитивнее по структуре, чем у полуденных песчанок. Группировки их менее прочны, а структура групп изменчива и сильно зависит от внешних факторов. В условиях лаборатории проявляется высокий уровень их агрессивности и социальной напряженности в группе. Однако эта особенность поведения у них сочетается со значительно меньшей стереотипностью выполнения движений при контактах, в частности сигнализирующих угрозу. У полуденных песчанок, в отличие от них, отсутствуют прочные социальные связи, в естественных условиях животные занимают индивидуальные участки. Их поведение в лаборатории высоколабильно и характеризуется отсутствием стереотипных ФКД. Агрессивные контакты этого вида очень интенсивны, но характерной чертой является незначительное влияние конфликта на поведение партнера.
Детальное исследование снятых в эксперименте на видеопленку эпизодов агрессивных контактов песчанок разных видов позволило выявить межвидовые особенности в организации видоспецифических движений, которые были незаметны при обычном визуальном анализе и которые могут служить примером анализа поведения близкородственных видов.
Разнообразие способов организации сообществ песчанок, которые мы рассмотрели выше, а также анализ социального поведения ряда других видов животных привели М.Е.Гольцмана (1983) к ревизии концепции доминирования с учетом сложных социальных ролей животных.
Голые землекопы (Heterocephalus glaber) — это восточноафриканские грызуны, которые роют многокилометровые сети подземных ходов в очень твердой почве. Эти животные (рис. 5.26) близки к грызунам подотряда Hystricomorpha, включающего также морских свинок и дикобразов. В 1981 г. Дж.Джарвис обнаружила, что они, подобно многим видам муравьев, живут колониями с единственной плодовитой самкой. Сходство с муравьями усиливается тем, что запутанная система туннелей и гнездовых камер колонии создается на основе совместной деятельности ее членов.
При рытье нор зверьки выстраиваются "конвейером", в котором переднее животное выполняет роль "проходчика", а далее следуют "откатчики" и "переносчики почвы". Они прорывают эти длинные туннели наугад, в разных направлениях, обеспечивая тем самым отыскание пищи — сочных клубней многолетних растений. Колонии голых землекопов насчитывают до 250 животных, у других видов млекопитающих такие крупные, сложно интегрированные группировки не известны. Наблюдения в лаборатории позволили выявить еще одну удивительную черту этих животных, которую до сих пор наблюдали только у общественных насекомых. Феномен заключается в том, что специализация зверьков меняется в зависимости от возраста и размеров. Молодые животные (в возрасте 2—3 месяцев), специализируются на "обслуживающих" операциях. Позднее эти же зверьки становятся фуражирами, а затем, вырастая, могут быть "охранниками", выполнять работу по выбрасыванию грунта или даже начать размножаться. Самый удивительный аспект жизни голых землекопов — особенности их размножения. Как и у многих видов общественных перепончатокрылых, у большинства самок этих грызунов репродуктивные функции подавлены, однако в отличие от насекомых самки грызунов не стерильны.
Если матка погибает, ее место занимает одна из рабочих самок. Когда самка-работница преобразуется в производительницу, у нее меняются и поведение, и внешний вид. Тело вытягивается за счет удлинения позвонков. Став маткой, такая самка приобретает способность подавлять половую активность других самок, а сама может приносить до 5 пометов в год, многократно спариваясь с разными самцами. Матка может также заставить "лентяев" работать, причем тем более активно, чем меньше зверьков действительно работает в данный момент (Reeve, 1992).
Этот тип организации сообществ (эусоциальность) ранее был описан только у общественных насекомых (см.: 5.6.2 и 5.7). Таким образом, даже у сравнительно примитивных млекопитающих — представителей отряда грызунов — типы социальной организации достаточно разнообразны.

Рис. 5.26. Голые землекопы
Подобно представителям многих других таксономических групп для разных видов хищных характерен широкий спектр типов организации сообществ как по сложности состава, так и по характеру взаимоотношений входящих в них особей. Среди них можно выделить таксоны с более консервативным типом социальной структуры (например, куницы и кошки) и таксоны с тенденцией к широкой радиации социальных систем (например, псовые). Рассмотрим особенности общественного поведения некоторых из них.
Бурый медведь — один из наиболее крупных хищников, особенности экологии и поведения которого позволяют ему успешно сущестовать даже на территориях, интенсивно осваиваемых человеком. В естественных условиях обитания бурый медведь — одиночно живущий зверь.
Внутривидовые отношения этого вида сводятся к кратковременному (несколько дней) контакту самца с самкой в брачный период. Кроме того, у медведя существует так называемая семейная группа, которая состоит из медведицы и двух—трех детенышей, родившихся зимой. Особенности поведения молодых и взрослых бурых медведей в течение более 20 лет исследовал B.C.Пажетнов (1985, 1991). Многолетние наблюдения позволили этому ученому описать закономерности формирования пищевого, исследовательского и оборонительного поведения у медвежат первого года жизни и использовать эти знания для разработки нового (приоритетного) метода восстановления численности этого вида. Межвежата-сироты поступают к B.C.Пажетнову, как правило, совершенно беспомощными, зимой, в период, когда проходит традиционная охота на берлоге.
Убивая медведицу, охотники, как правило, забирают медвежат и выращивают их дома. Судьба таких животных предрешена: они не боятся человека, делаются "ручными" и в течение нескольких месяцев радуют своих владельцев. Постепенно звери становятся опасными, и их либо передают в зоопарки и цирки, либо убивают. Работа Пажетнова показала принципиальную и практическую возможность вырастить таких межвежат "в боязни перед человеком", что позволяет успешно выпускать их в дикую природу: у них сохраняется устойчивая реакция избегания человека и следов его жизнедеятельности. После начального периода выкармливания медвежат из соски, начиная с возраста 2,5—3 месяца медвежат оставляют одних в изолированном от шума помещении, приносят им корм один раз в сутки. Человек, с которым медвежата контактируют наиболее часто, воспринимается ими в роли матери: они следуют за ним по пятам с первых дней самостоятельного передвижения по лесу. Это так называемая реакция следования, формирование которой, видимо, происходит по типу запечатления (см.: 7.1). Медвежата неотступно следуют за человеком в течение многокилометровых экскурсий по лесу, длящихся иногда несколько дней. По мнению B.C. Пажетнова, такая реакция следования обеспечивает устойчивость семейной группы бурого медведя.
Наблюдения за формированием пищевого поведения позволили уточнить список видов растений, наиболее часто используемых детенышами, а также установить преобладайте источники основных пищевых ингредиентов. К концу июня пищевые навыки молодых медвежат уже сформированы, и наступает фаза формирования оборонительного поведения. Это выражается в появлении сильного испуга в ответ на все новые раздражители. До этого медвежата также могли пугаться каких-либо предметов или ситуаций, но реакция испуга бывала кратковременной.
Начиная с конца июня, т.е. с возраста примерно 4,5—5 месяцев, реакция страха делается интенсивной и устойчивой. В ноябре, после накопления жира такие медвежата готовятся к зимнему сну, и Пажетнову удалось увидеть последовательные фазы такой подготовки, начиная с выбора места для берлоги. Ни предшествующего опыта строительства, ни примера для подражания у этих животных не было. Записанные на видеопленку действия медведей поражают четкостью и целесообразностью. В ходе строительства медвежата-сеголетки иногда по каким-то причинам бросают начатую берлогу и переходят в новое место. На основании многолетних наблюдений автор приходит к выводу, что для выполнения такого сложного действия, как строительство берлоги, "жизненного опыта" не требуется.
У перезимовавших медвежат сохраняется реакция избегания людей, и по поведению они практически не отличаются от диких сородичей. Удалось установить размеры территории, которыми "владеют" самцы и самки медведей разных возрастных категорий, и определить влияние антропогенного фактора на численность популяций. На основе этих наблюдений разработаны и успешно применяются методы воспитания медвежат-сирот и возвращения их в природу. Первый опыт выращивания медвежат-сирот описан в популярной книге Пажетнова "Мои друзья-медведи"(1985), а научное описание работы с ними и результаты наблюдений он обобщил в монографии "Бурый медведь" (1991).
Волк. Поведение волков, которые относятся к другому семейству отряда хищных (псовые), исследовалось как в природных условиях, с использованием традиционных методов тропления и визуального наблюдения (Бибиков, 1985), так и у прирученных (Бадридзе, 1987) и полуприрученных особей (Крайслер, 1966). Волки живут семейными группами в период размножения и выращивания молодняка и объединяются в стаи в осенне-зимний сезон. Стая волков — это закрытое индивидуализированное сообщество, оно характеризуется наличием четкой иерархической структуры, которая поддерживается с помощью разнообразных агрессивных контактов. Члены стаи умеют четко согласовывать свое поведение в таких сложных коллективных действиях, как, например, охота на лося. Гиеновые собаки. Примером стабильного закрытого индивидуализированного сообщества хищных млекопитающих, основанного на устойчивых дружеских связях между его членами, могут служить гаеновые собаки (Lycaon pictus), очерк поведения которых дали Дж. и Г.Лавик-Гудолл в книге "Невинные убийцы" (1977; см. также: Крушинский, 1986. С. 184—185). Стая, насчитывающая до 40 животных, ведет кочевой образ жизни в пределах большого участка (до 4000 кв. км). Роль лидера в ней может принадлежать животным обоих полов, однако чаще это бывают самки. В период размножения щенится обычно только самка с наиболее высоким иерархическим статусом. Пары, включающие самок более низкого ранга, временно покидают группу, Первые три недели после родов, когда самка остается со своими щенками, члены группы коллективно охотятся и регулярно кормят ее и детенышей. Кооперация и взаимная социальная терпимость могут сочетаться в стае с коллективным деспотизмом, при котором, однако, низкоранговые особи не выселяются за пределы группы.
Сообщества гиеновых собак организованы по-другому, чем сообщества волков. Проявления агрессии играют относительно небольшую роль в регуляции внутригрупповых отношений. Драки крайне редки, основную роль играют ритуализованные действия. При встрече двух стай происходят агрессивные столкновения, которые препятствуют их смешиванию. Тем не менее, ведущую роль в организации стай играет общественный инстинкт, который выражается в совместной охоте, выкармливании потомства и т.д.
Низшие приматы. Сообщества низших приматов имеют разнообразную структуру. Среди них можно найти виды, которые живут относительно небольшими семейными группами, а также виды, сообщества которых напоминают сообщества псовых. В 60—70-е годы этими вопросами занимались многие исследователи.
Полевые наблюдения за общественными отношениями в группах этих приматов легли в основу современных представлений об общественном поведении животных. Работы Дж.Крука (Crook, 1970a, б; 1977), И.ДеВоре (De Vore, 1965 а, б) и др. выявили многообразие, видоспецифичность, а также определенный уровень лабильности группировок низших обезьян (см. также: Хайнд, 1975; Меннинг, 1982). Выше говорилось о многих особенностях их oбщественного поведения.
Так, рис. 5.13 показывает структуру стаи павианов при пересечении ими открытого пространства. Низшие приматы образуют, как правило, индивидуализированные сообщества закрытого типа, в которых его членам обеспечена безопасность от внешних вмешательств. В этих сообществах поддерживается система ранговой иерархии, которая, однако, не исчерпывает всей сложности их взаимоотношений. Благодаря высокому уровню рассудочной деятельности они способны реагировать не только на сигналы угрозы и подчинения, но улавливают и более тонкие особенности взаимоотношений между членами группы. Отмечены случаи, когда обезьяны используют безусловное "табу" на действия, которые могут причинить вред детенышам, для предотвращения или решения конфликтов. На рис. 5.14 показано, как молодой самец берберийского макака демонстрирует высокоранговому самцу свой тесный контакт с детенышем, чтобы добиться его благосклонности (см.: Меннинг, 1982). У многих видов развита взаимопомощь (кооперация) в разных сферах деятельности.
Классическое описание социальной структуры сообщества павианов-гелад и ее динамических изменений можно найти в статье английского этолога Дж.Крука (1977), подробно они изложены в книге Л.В.Крушинского (1986. С. 199—204). Особенно важно, что Дж.Крук одним из первых обратил внимание на огромную роль в общественных отношениях обезьян не только конкуренции, но и взаимопомощи, сотрудничества. Он выделил две формы взаимопомощи. Первая — "содействие" (cooperation), т.е. такое поведение группы животных, которое направлено на выполнение определенного преднамеренного действия, имеющего общую выгоду (например, удовлетворение эмоциональных потребностей или получение "награды" для всех членов сообщества). Другая форма взаимодействия — "сотрудничество" (collaboration) — разделение между членами сообщества отдельных фрагментов выполняемых действий. Обе формы взаимоотношений удается наблюдать у птиц и некоторых видов млекопитающих при создании ими пищевых запасов на зиму, защите обитаемой территории, взаимной защите от хищников, совместной охоте, заботе о потомстве и т.д. У обезьян взаимодействие выражено наиболее ярко.
Сотрудничество у мартышковых обезьян проявляется, например, при заботе о новорожденных детенышах, которую берет на себя не только мать, но и одна из дружественных матери бездетных самок. По-видимому, в естественных условиях жизни забота бездетных самок о детенышах как при жизни, так и в случае смерти матери имеет большое биологическое значение. У некоторых видов мартышковых обезьян (например, макак) популяция разбивается на несколько групп, которые можно назвать "кастами". Есть основания считать, что "подруги" матерей, берущие на себя заботу о сироте в случае смерти самки, — это ее "близкие родственники". Помогая матери или беря на себя заботу о детенышах в случае ее гибели, они способствуют сохранению генотипа своей "касты". Такая форма "альтруистического" поведения развилась, несомненно, в результате группового отбора, на возможность которого указывал Ч.Дарвин. Те касты родственных животных, которые несли в своем генотипе предпосылки для выполнения описываемой формы "альтруистического" поведения, имели шансы вырастить большее число детенышей, чем касты, не обладавшие подобным генотипом. Поэтому селекция на альтруизм закрепляет определенные нормы поведения в группе родственных обезьян, обеспечивая сохранение присущего ей генотипа. В ряде случаев подобную заботу о потомстве могут проявлять и самцы.
Самцы диких японских макаков (Масаса fuscata) ухаживают за детенышами, защищают их от других обезьян и всевозможных врагов. Покровительство детенышам оказывают в основном самцы, находящиеся на среднем уровне иерархических отношений. Предполагается, что те обезьяны, которые ухаживают за детенышами доминантных особей, повышают свое положение на иерархической лестнице. Как отмечает Л.В.Крушинский, обнаруженные этими исследователями факты привлекли внимание к сложности отношений, устанавливающихся между членами колоний мартышковых обезьян. Во всяком случае, они гораздо сложнее, чем иерархические отношения, складывающиеся на основе агрессии в группах других животных. Они определяются не только способностью к бою, но и способностью найти поддержку со стороны других самцов. На этом основании было сформулировано понятие "роли", которое базируется на динамике многообразных взаимоотношений в сообществах животных (см. также: 5.3.7). В каждой группе животных отдельные особи реагируют специфически на других членов этого сообщества, например на самцов, самок, доминантных, подчиненных им особей и т.д. Совокупность социальных контактов каждого животного определяет его "роль" в сообществе. Таким образом, в сообществе мартышковых обезьян взаимопомощь следует рассматривать как существенный фактор, обеспечивающий выживание родственных особей и достижение ими определенного положения в иерархической структуре.
Л.В.Крушинский высказал предположение, что в динамике сложных взаимоотношений в сообществах мартышковых обезьян их высокоразвитая элементарная рассудочная деятельность должна играть существенную, а, возможно, и ведущую роль. Это представление получило подтверждение и развитие в работах Гудолл (1992) на шимпанзе.
Шимпанзе. Выше неоднократно приводилось описание отдельных сторон социального поведения шимпанзе. Из всех видов млекопитающих они характеризуются наиболее удивительной социальной структурой. Их сообщества, состоящие из 50 или более членов, занимают территорию, с которой изгоняются все другие самцы шимпанзе.
В разделе 5.2.4 при описании особенностей индивидуализированных сообществ рассматривалась специфика тех социальных контактов, с помощью которых поддерживается сложная структура сообществ шимпанзе и других человекообразных обезьян. Эти и многие другие, практически исчерпывающие сведения о структуре сообществ шимпанзе, были получены Дж.Гудолл благодаря непрерывному наблюдению за популяцией этих обезьян в Танзании, которые проводились под ее руководством в течение нескольких десятилетий. Они подробно изложены и обобщены в монографии "Шимпанзе в природе: поведение" (1992), которая освещает практически все стороны поведения этих животных: демографические особенности, использование территории и связанные с этим реакции, питание, охоту, манипуляционное и половое поведение, отношения с обезьянами других видов (главным образом с павианами), биографии отдельных его членов.
Социальному поведению посвящена значительная часть этой книги. Гудолл рассматривает природу сообществ шимпанзе и подробно анализирует его уникальные признаки. Один из таких признаков — характерная для шимпанзе вариабельность социальных контактов. Шимпанзе держатся группами, хотя известно, что они неплохо существуют в одиночку. Более того, одиночки и малые группы (до 3—4 особей) при поиске пищи нередко оказываются в более выгодном положении. Тем не менее, как было установлено, самцы специальными звуковыми сигналами ("уханием") оповещают членов сообщества о найденной пище и привлекают их к плодовым деревьям. Подобную кооперацию самцов шимпанзе некоторые авторы объясняют с точки зрения концепции "отбора родичей" (см.: 6.5.1).
В предыдущих разделах было показано, что у представителей разных таксономических групп позвоночных социальные структуры имеют принципиальное сходство. Это послужило основой для формирования представления об изоморфизме типов сообществ (см.: 5.7). Как известно, строение нервной системы у беспозвоночных и позвоночных принципиально различно. Поэтому вопрос о типах социальных структур у беспозвоночных приобретает особый интерес. Рассмотрим, насколько принцип изоморфизма распространяется на сообщества беспозвоночных.
Наряду с многочисленными одиночными видами, среди беспозвоночных обнаружены и виды, объединяющиеся в сообщества разных типов.
Многие виды беспозвоночных образуют скопления, или агрегации (объединения), под влиянием тех или иных факторов среды. Это, например, скопления планктонных рачков в местах, богатых пищей, или миграционные скопления стадных саранчовых.
Такие группы сообществами не являются, в них отсутствует упорядоченная структура, однако особи в них реагируют на присутствие друг друга — это так называемый эффект группы. У рачков он выражен слабо и проявляется в увеличении скорости питания в зависимости от размера группы. В то же время у перелетной саранчи, как это было впервые установлено в 1926 г. Б.П.Уваровым, происходят глубокие физиологические изменения. Зеленые саранчуки, находясь в группе, изменяют цвет и превращаются в черно-красных, изменяется и форма их тела: они становятся "горбатыми". Коренным образом меняется поведение: у них явно выражено стремление к объединению в огромные стаи и миграции. В этом примере видна некоторая условность приведенной выше (см.: 5.2.2) классификации сообществ, поскольку такие стаи, помимо черт, характерных для агрегаций, обладают рядом признаков анонимных сообществ открытого типа.
Индивидуализированные сообщества, в которых особи узнают друг друга и отличают от посторонних, образуют не только общественные насекомые, но и другие членистоногие. Например, у пустынных мокриц подбор пар осуществляется путем длительных конфликтов, однако затем пары сохраняются, и насекомые строят свои норки вдвоем, охраняя и опознавая появившееся потомство.
Отличительной чертой социального поведения насекомых некоторых видов (их называют общественными) является наличие у них "эусоциальности" (т.е. истинной социальности). Этим термином обозначают наиболее сложную форму общественной организации, которая, как считалось до недавнего времени, характерна только для насекомых.
У таких видов социальная организация удовлетворяет следующим критериям.
Во-первых, в группе существует так называемая репродуктивная дифференциация, или разделение репродуктивных функций: одни особи размножаются, а другие — рабочие особи (их, как правило, большинство) — бесплодны и выкармливают потомство плодовитых особей. У муравьев, например, все население муравейника, кроме скрытых в глубине самок ("цариц") и появляющихся раз или дважды в год мелких крылатых самцов, представляет собой бесплодных рабочих самок. Часть из них приносит корм (фуражиры), часть поддерживает в муравейнике чистоту, кормит личинок, охраняет и т.д. (подробнее см. далее).
Во-вторых, между членами группы наблюдается кооперация, т.е. совместное добывание пищи, выкармливание потомства, строительство, защита гнезда и т.п.
В-третьих, особи не менее двух последовательных поколений — материнского и дочернего — живут вместе на тех стадиях жизненного цикла, когда они выполняют функцию "рабочих".
Сходным образом организованы сообщества других общественных перепончатокрылых (ос, пчел, шмелей), а также термитов. Кроме того, эусоциальность обнаружена у ряда японских тлей и у одного вида австралийских жуков. В начале 80-х годов аналогичная система была открыта у позвоночных — у так называемых "голых землекопов" (см.: 5.5.1).
В большинстве случаев узнавание беспозвоночными друг друга аналогично узнаванию членов своей колонии у грызунов, где ключевую роль играет специфический запах колонии и/или особи. В то же время установлено, что общественные насекомые, например муравьи многих видов, не просто опознают членов своей семьи по принципу "свой—чужой", но действуют на своем кормовом участке в составе небольших групп, знакомых друг с другом непосредственно (см.: 5.6.4).
Появление в эволюции насекомых эусоциальности заставляло ученых искать промежуточные формы организации поведения в группах, т.е. такие формы, которые еще нельзя назвать сообществами, но которые уже отличаются от одиночного образа жизни и от агрегаций. Как следует из сравнительных исследований, эволюция эусоциальности, по-видимому, причинно связана с репродуктивной стратегией вида (см.: 6.5.1). Еще в 20-е годы исследователи смогли выделить 7 этапов развития материнского поведения, которое охватывает все уровни социальных структур в пределах класса насекомых. Эта последовательность, по всей видимости, описывает практически все типы общественных структур насекомых:
1) самка беспорядочно рассеивает яйца (например, поденки, некоторые жуки);
2) самка откладывает яйца вблизи источника пищи или непосредственно на пищу;
3) самка снабжает отложенные яйца защитной оболочкой (пушком, воскоподобными выделениями) или укрытием;
4) самка остается на некоторое время с яйцами или молодыми личинками и охраняет их (например, уховертки, некоторые сверчки, клопы-щитники);
5) самка откладывает яйца в специальное укрытие или специально вырытое гнездо и снабжает их запасом провизии (жуки-навозники, одиночные пчелы и осы; последних помещают в запечатываемые ячейки парализованных насекомых);
6) самка остается со своими личинками в гнезде, защищает их и снабжает пищей (например, осы-бембексы, гнездящиеся в норках на открытых песчаных участках, выкармливают своих личинок, принося им ежедневно несколько убитых мух);
7) потомство остается в гнезде с матерью и совместно с ней выращивает следующие поколения особей.
На этом последнем этапе появляются предпосылки для возникновения эусоциальности. Своего наивысшего расцвета среди насекомых эта система организации сообществ достигла в двух систематически далеких группах — у равнокрылых (термиты) и у жалящих перепончатокрылых (муравьи, пчелы, осы). Возможные пути эволюционных преобразований структуры сообществ, в том числе и у беспозвоночных, рассматриваются далее (см.: 6.5.1).
К настоящему времени накоплено множество сведений о социальном поведении насекомых на основании наблюдений за ними как в естественных, так и в лабораторных условиях, где можно видеть все происходящее в улье с прозрачными стенками или в формикарии — искусственном прозрачном муравейнике. Формикарии размещают на больших лабораторных аренах, позволяющих наблюдать любые контакты между насекомыми. Индивидуальная цветная метка дает возможность знать муравьев "в лицо". Метки могут быть групповыми или индивидуальными и наносятся на разные части тела в виде разноцветных точек.
Рассмотрим основные черты организации сообществ у эусоциальных насекомых.
Термиты. Это довольно примитивные насекомые, но они образуют огромные и высокодифференцированные сообщества. Термиты — главные преобразователи веществ в почве от тропиков до пустынь: они разрыхляют и перемешивают почву и с помощью бактерий, простейших и грибов разрушают волокна целлюлозы, возвращая их компоненты в общий круговорот веществ. Найденную целлюлозу они переносят в запечатанные холмики с постоянными условиями температуры и влажности, где она разлагается с выделением энергии. Разновозрастные личинки выполняют в семье различные работы. В семействах термитов полиморфизм таких бесполых, ювенильных особей выражен по-разному. Группы с морфологически закрепленными особенностями, связанными с различной ролью в семье, называют кастами. Особенно замечательна у некоторых видов термитов каста солдат: у одного вида, получившего название "носатых термитов", это "носатые" солдаты с выростом на голове, с помощью которого они выбрызгивают струю отпугивающей жидкости, у другого солдаты имеют мощные защелкивающиеся мандибулы (челюсти).
Муравьи. примере муравьев рассмотрим более подробно устройство семьи общественных насекомых. Муравьи, пчелы и осы произошли от более высокоорганизованных, чем термиты, насекомых с более развитыми органами чувств, которые не боялись света. Эти насекомые распространены чрезвычайно широко и оказывают существенное воздействие на экосистемы суши, выступая в роли охотников, мусорщиков, потребителей углеводных выделений насекомых и растений, опылителей и переносчиков семян. Их сообщества могут быть крошечными и огромными, лабильными или стационарными. Из названных трех групп насекомых муравьи представлены одними лишь общественными видами, а среди ос и пчел много одиночных видов. У всех общественных перепончатокрылых семьи полиморфны: помимо самцов и самок, они имеют многочисленных рабочих, неполовозрелых самок, выполняющих работу по строительству гнезда и заготовке провизии, выращиванию потомства и кормлению продуцирующей яйца самки (их называют также матками, или царицами). У пчел, шмелей и цветочных ос потомство выкармливается медом и пыльцой, у общественных ос — убитыми насекомыми, у муравьев — белками животного и растительного происхождения.
Поскольку муравьи, как и термиты, происходят от видов, первоначально обитавших в почве, у них есть черты экологического и морфологического сходства, несмотря на то, что в систематическом плане равнокрылые и перепончатокрылые весьма далеки: как и у термитов, у муравьев самки обламывают свои крылья (у термитов — до копуляции, у муравьев — после), а рабочие особи бескрылы от рождения. Как и у термитов, рабочие у муравьев могут принадлежать к различным кастам, но если у термитов рабочие — это личинки разных возрастов, причем как самцов, так и самок, то у муравьев все рабочие (как у пчел и ос) — это взрослые, но неполовозрелые самки. У некоторых видов муравьев принадлежность к разным кастам закреплена морфологически, и члены семьи различаются по размерам тела, размерам и форме головы, что соответствует их функциям в семье: так, у одного из видов муравьев-древоточцев есть каста "затычек", члены которой предназначены для закрывания головой входов в гнездо, устроенное в дереве. У некоторых видов границы каст весьма условны и различаются только размерами.
Касты дифференцируются в личиночном возрасте, и этот сложный процесс определяется не только наследственной программой, но и составом пищи, различными феромонами и своеобразным массажем (за счет сдавливания челюстей), который производят рабочие, ухаживающие за молодью. Поведенческие стереотипы рабочих и соответственно их функции в семье определяются либо принадлежностью к определенной касте, либо меняются с возрастом. Последняя форма "разделения труда" между особями разных возрастов называется "возрастным полиэтизмом" и описана у медоносной пчелы и некоторых видов муравьев: особь переходит от внутригнездовой деятельности к различным видам работ вне гнезда.
Удивительно слаженное функционирование семей общественных насекомых основано на разделении труда между группами особей, и обычно чем крупнее семья, тем более четко выражена специализация. Так, в большом муравейнике одни муравьи ухаживают за размножающимися царицами и молодью, другие занимаются строительством, третьи снабжают население гнезда углеводной пищей (сладкими выделениями различных насекомых, соком растений), четвертые приносят белковую пищу (например, живых или мертвых беспозвоночных), пятые заняты охраной границ или выполняют функцию наблюдателей. На примере муравьев достаточно четко прослежена зависимость уровней социальной организации от численности семьи. У многих видов муравьев (а их несколько тысяч) можно выделить ряд уровней организации. Так, при численности семьи в несколько сотен особей, муравьи живут в сравнительно просто устроенных подземных гнездах и не охраняют свой участок. Отдельные фуражиры действуют самостоятельно, они приносят в гнездо мелких насекомых (для выкармливания личинок) и транспортируют в своих зобиках углеводную пищу. Внутри гнезда эта пища распределяется между взрослыми муравьями. При других уровнях социальности в семье муравьев действует такая же система жизнеобеспечения, однако при этом и организация семьи, и структура территории иные. Например, при численности семьи порядка сотни тысяч особей муравьи строят то, что мы называем "муравейником".
Муравейник — это гнездо из сухих травинок и веточек, самая сложная часть которого находится под землей. У таких семей кормовая территория охраняется, она пронизана сетью постоянных фуражировочных дорог, ведущих к деревьям, кустам или травянистым растениям с колониями тлей, каждая дорога обслуживается определенной группой муравьев. Такие малые группы постоянны по составу и хорошо различают друг друга. Муравьи в таких семьях склонны к кооперативным действиям. На территории, принадлежащей муравейнику, можно наблюдать групповую транспортировку пищи и даже групповую охоту. При миллионном населении муравейники начинают "почковаться" — давать "отводки" путем выселения части муравьев, "цариц" и личинок на новое место. Так возникают колонии — группы родственных гнезд, связанных обменными дорогами. Объединение колоний, занимающее иногда целые лесные массивы, называют федерацией.
Такова общая схема функционирования семьи муравьев как организованного сообщества эусоциальных насекомых. Более подробно о социальной жизни муравьев можно прочитать в книге А.А. Захарова "Муравей, семья, колония" (1991). Общественное устройство колонии медоносной пчелы описано К. Фришем (1980) и Н.Г. Лопатиной (1971).
Индивидуализированные группы в семье. Как отмечалось выше, в семье общественных насекомых выделяют так называемые функциональные группы. У медоносной пчелы это сборщицы пыльцы и нектара, разведчицы (они же "танцовщицы"), "няньки", ухаживающие за личинками. У муравьев выделяются активные фуражиры, занятые в основном охотой, трофобионты ("живые вагончики"), добывающие и транспортирующие в гнездо жидкую углеводную пищу, внутригнездовые рабочие и некоторые другие группы.
Взаимодействие особей в таких группах большинством авторов рассматривалось как анонимное. Возможно, что так оно и есть для многих видов общественных ос и шмелей. Так, А.Ю.Дудников (1983) показал, что у ос-веспин индивидуальное взаимодействие регулируется иерархией мотиваций, не требующей индивидуального узнавания: при конфликте на источнике пиши побеждает та оса, которая более голодна в данный момент.
Разделение труда в семье основано на том, что каждая оса в течение жизни по очереди выполняет разные роли. В то же время есть указания на то, что у одного из видов общественных ос (Metapolibia cingulata) взаимодействие между особями строится на основе доминирования-подчинения, и попадая в чужую семью, оса теряет ранг (Forsyth, 1975).
Сходный феномен был открыт А.А.Захаровым (1991) на рыжих лесных муравьях. Этот автор описал индивидуализированные группы в семье муравьев — кланы, в которых лидер первым атакует добычу, нередко отнимает ее у других муравьев, часто вступает в схватки с муравьями, близкими по рангу.
Выяснение иерархического ранга особи в группе часто сопровождается своеобразными турнирами, когда фуражиры пытаются унести друг друга в гнездо, несмотря на обоюдное сопротивление. Побежденный муравей принимает специфическую позу (складывается "чемоданчиком") и позволяет себя транспортировать.
Такое поведение является одним из примеров ритуализованных действий муравьев, удивительно сходных в этом плане с поведением позвоночных животных. Так, в ответ на агрессивные выпады муравей предлагает партнеру пищу, что рассматривается как демонстрация покорности. Медовые муравьи, встретив чужаков, подолгу стоят попарно в угрожающих позах, удивительно напоминающих позы собак: ноги выпрямлены и напряжены, головы повернуты в стороны друг от друга (Holldobler, 1976). Луговые муравьи-"пограничники" из разных семей, встречаясь регулярно на границах территорий, опознают друг друга и ограничиваются демонстрациями угроз, тогда как незнакомых "пограничников" незамедлительно уничтожают (Резникова, 1979). Чтобы выяснить это, пришлось привязать муравьев на тонкие поводки, как собачек, прикрепив поводки к предметным стеклам с нанесенным на них шифром. На границе муравьиной территории были расставлены наборы стекол, к которым были привязаны муравьи, взятые из той же семьи (контрольные), чужие, но все же знакомые муравьи-"пограничники", а также совсем незнакомые муравьи, взятые из глубинной части территории соседней семьи. Муравьи на поводках вели себя достаточно спокойно и активно, а поведение "пограничников" изменялось от полного нейтралитета до безоговорочного уничтожения.
Детальные наблюдения в природе за работой индивидуально помеченных муравьев выявляют удивительно тонкую дифференциацию ролей в маленьких, слаженно работающих группах. В лабораторных экспериментах, посвященных сложным формам коммуникации муравьев (см.: 5.4.5), было показано, что в пределах такой "малой рабочей группы", насчитывающей около десятка особей, происходят сложнейшие процессы передачи информации, в том числе с использованием дистанционного наведения, когда муравей-разведчик, вступая в контакт с фуражирами, передает им сведения о координатах кормушки, которую они затем находят самостоятельно, не пользуясь пахучим следом. Такие группы могут не только искать пищу, но и решать возникающие при этом задачи, такие как избегание опасности или преодоление препятствий.
Так, в одном из экспериментов группы муравьев вместе с личинками и коконами помещали в изогнутую трубку. Чтобы попасть в затемненную часть трубки и там спрятать расплод от света, муравьи должны были вытащить затычки из перегородки, закрывающей им путь. Оказалось, что с такой задачей способны справиться лишь хорошо "сработавшиеся" группы, причем одни муравьи вытаскивали затычки, а другие переносили расплод (Резникова, Новгородова, 1998).
Все перечисленные факты свидетельствуют в пользу точки зрения о гибкости и сложности индивидуального поведения общественных перепончатокрылых и о значительной роли отдельной особи в принятии решений, жизненно важных для семьи. Вполне возможно, что именно на пластичности поведения отдельных животных основаны интегрированные реакции семьи в целом. В качестве одного из примеров можно привести поведение семьи муравьев рода Formica, подвергавшихся в эксперименте постоянному гамма-облучению. В ответ на появление этого фактора муравьи выстроили крытую дорогу длиной 12 м, что позволило им уменьшить дозу радиации (Brower, 1966). В экспериментах Ж.И. Резниковой (1979) было показано, что муравьи одного вида могут быстро менять уровень социальной организации, в буквальном смысле на глазах экспериментатора. В естественной ситуации у муравейника лугового муравья (Formica pratensis) были огорожены участки фуражировочных дорог, что вынуждало муравьев построить обходные дороги и перераспределить активность на внутренних частях кормового участка. В конечном итоге в одном из секторов кормового участка этой семьи динамическая плотность муравьев повысилась в несколько раз.
Поразительна была скорость изменений поведения целой семьи. В течение одного месяца муравьи построили вспомогательные гнезда, полностью изменили в связи с этим систему фуражировки на территории, затем в одно из этих вспомогательных гнезд была принята оплодотворенная "царица". Вскоре там появились личинки и в конце концов образовалась "локальная колония" со своей обменной дорогой. Таким образом, поведение муравьев существенно усложнилось. Можно сказать, что семья перешла от уровня одиночной охраняемой территории к уровню колонии. В опытах с другими видами муравьев экспериментаторы воздействовали на численность семьи, подкладывая муравьям в гнездо "приемышей" — куколки собственного вида, из которых вскоре выходили новые муравьи (Резникова, 1983). Действуя достаточно деликатно, можно было довести число "приемышей" до нескольких тысяч. Так удавалось заставить муравьев, изначально имевших простое подземное гнездо и неохраняемый, "бесструктурный", кормовой участок, строить земляной купол. У них появлялся аналог типичной фуражировочной дороги и групповая транспортировка добычи.
Эти результаты интересны тем, что выявляют удивительную пластичность социального поведения одних и тех же животных, быстро перестраивающегося под воздействием определенных факторов, т.е. свидетельствуют о высокой пластичности индивидуального поведения отдельного муравья или пчелы.
Исследование общественного поведения огромного числа видов животных разных таксономических групп позволило выявить уже упоминавшийся выше феномен так называемого изоморфизма, структурного сходства социальных систем в различных, не связанных между собой таксонах.
Эусоциальность, считавшаяся феноменом, свойственным только насекомым, была обнаружена, как об этом говорилось в разделе 5.5.1, у млекопитающих. При этом налицо и противоположное явление: у относительно близкородственных видов, подчас относящихся к одному и тому же роду, внутрипопуляционная организация нередко различается значительно сильнее, чем у видов, принадлежащих к филогенетически удаленным друг от друга таксонам. Столь высокую степень сходства социальной организации у разных видов обычно нельзя объяснить явлением конвергенции, так как оно встречается у видов, занимающих разные экологические ниши.
Общей чертой общественного устройства животных разных таксономических групп является феномен иерархических рангов, в установлении которых может обнаруживаться большое разнообразие, связанное с видовой и экологической спецификой данной группы.
Выше были приведены примеры ряда социальных систем, характерных для разных видов животных во многих экологических ситуациях, а также известные способы поддержания их структуры. Эти примеры показывают, что роль особи в группе может зависеть от ее пола, возраста, физиологического состояния, индивидуального и социального опыта, а также сочетания всех этих качеств с конкретной ситуацией.
Описание общественного поведения животных — тема для большой монографии. Очевидно, что многие вопросы этой огромной темы в данной главе были лишь затронуты. Относительно большее внимание мы уделили тем способам коммуникации, которые требуют от них высокого уровня когнитивных способностей животных. Эти данные относительно новы и на русском языке в нашей книге они описываются практически впервые.