
Первые организмы
Самые первые системы, способные эволюционировать под действием естественного отбора, видимо, были устроены иначе, чем современные организмы, и имели иной состав. Ими могли быть кристаллы глины
А.ДЖ.КЕРНС-СМИТ
ПРИ ВСЕМ разнообразии ныне существующих на Земле форм жизни у них есть общее: главные молекулярные механизмы у современных организмов, как было неоднократно показано, одни и те же. Установление этой общности биохимической организации — одно из крупнейших открытий за последние сто лет. Нет никакого сомнения, что оно проливает свет и на историю эволюции. Но, как мне кажется, при рассмотрении самых первых этапов эволюции концепция единства биохимической организации не приносит большой пользы.
Сказанное противоречит наиболее широко распространенной точке зрения относительно зарождения жизни. Принято считать, что до возникновения первых организмов, или, иными словами, до возникновения систем, способных к неопределенно долгой эволюции под действием естественного отбора, происходила иная, химическая эволюция, в ходе которой образовался некий набор молекул, своего рода универсальный конструктор, из которого «собраны» ныне существующие организмы. В этот набор входили аминокислоты, сахара и молекулы других типов.
Такая точка зрения имеет свои корни в исследованиях 20-х годов, когда советский биохимик А.И.Опарин и английский биолог Дж.Холдейн создали концепцию «первичного бульона», содержавшего органические молекулы, которые существовали в океанах до зарождения жизни на Земле. Предполагалось, что «бульон» образовался в результате геохимических процессов и воздействия различных энергетических процессов на атмосферу, которая в чем-то напоминала существующую сегодня на Юпитере — в ней преобладали неокисленные газы метан, аммиак и водород. Этого мнения относительно состава первичной атмосферы придерживался Г.Юри. В его пользу говорили результаты опытов С. Миллера (в ту пору студента в лаборатории Юри), которые он выполнил в начале 50-х годов. Через смесь газов, которые предположительно входили в состав примитивной атмосферы, Миллер пропускал электрические разряды («молнии») и после этого обнаружил в ней водорастворимые молекулы органических веществ. Не менее 15% углерода, входившего первоначально в состав метана, превращалось в относительно небольшое число некрупных молекул, включая четыре из двадцати аминокислот, образующих белки — эффект весьма впечатляющий. Не менее убедительны результаты экспериментов X.Оро, которые он провел в начале 60-х годов. Оро показал, что молекулы синильной кислоты HCN в одностадийной реакции могут конденсироваться с образованием аденина. Ранее Миллер продемонстрировал, что синильная кислота может образовываться в опытах с электрическими разрядами. При этом происходило образование малых молекул еще одного типа — формальдегида СН2О; а уже лет сто известно, что молекулы формальдегида также способны объединяться с образованием Сахаров, таких, как рибоза, являющаяся одним из компонентов РНК.
Думалось, что формирование адекватного «конструктора для начинающих» в таких условиях было лишь вопросом времени. Замечание скептиков, что первичная атмосфера на Земле могла иметь иной состав, чем ныне на Юпитере (а сегодня так считают многие), не казалось существенным, поскольку эксперименты с искусственными атмосферами другого состава и при использовании иных источников энергии давали похожие смеси аминокислот.
ВСЕ ЖЕ первоначальные предположения не нашли подтверждения. Экспериментальный подход, предложенный Миллером, не имел развития. В таких опытах при имитации условий, существовавших на ранних этапах развития Земли, даже простейшие молекулы образуются в очень небольших количествах. Дело осложняется и тем, что эти молекулы составляют обычно лишь малую часть продуктов реакций. Трудно представить себе, как осуществлялись их отделение и очистка в ходе геохимических процессов, в которых обычно смеси органических веществ, напротив, становятся все более беспорядочными. Чем сложнее молекулы, тем более усугубляются эти противоречия. В частности, трудно представить себе чисто геохимическое образование нуклеотидов (мономеров ДНК и РНК). Во всяком случае, в имитационных экспериментах типа тех, что ставил Миллер, образование нуклеотидов до сих пор не наблюдалось.
Остается ли, несмотря на все сказанное, в пределах здравого смысла предположение, что конструктор должен был образоваться в самом начале? Действительно или ложно, что:
1) самые главные молекулы жизни одинаковы у всех организмов, живущих сегодня на Земле;
2) по меньшей мере некоторые из этих молекул могли образоваться в условиях, характерных для ранних стадий истории Земли?
Я думаю, что эти утверждения уводят нас в сторону, а то, что их два, еще больше ухудшает дело.
Неправомерность первого из них выявляется при анализе единства биохимии, когда мы вспоминаем, что для всех организмов общим является нечто большее, чем конструктор из малых молекул. Имеется система — общий план организации, который всегда одинаков. Взаимодействие механизмов в такой системе чрезвычайно сложное. Даже единичная молекула белка — это сложное образование, в котором специфически соединены тысячи атомов, а ведь для жизнедеятельности нужны сотни таких должным образом организованных молекул. Одно из звеньев, где потребность в белках выражена наиболее ярко (и где они должны быть наиболее «noдогнаны»), — механизм биосинтеза белка. Этот типичный пример клубка проблем представляет другой аспект единства биохимии — тесную взаимосвязь всех компонентов жизнедеятельности. Наконец, существует и произвольность в наших представлениях относительно некоторых свойств самого комплекса основных механизмов. Так, код для «перевода» текста РНК в последовательность белка, всюду практически одинаков, а набор аминокислот везде один и тот же. Трудно поверить, что был возможен только один работоспособный код или один набор аминокислот или что они были наилучшими для всех организмов при всех обстоятельствах.
Вне сомнения, наиболее правильно заключить из этого детального рассмотрения единства биохимии следующее:
1) все живое на Земле происходит от общего предка,
2) на эволюционном древе этот предок располагается достаточно высоко и
3) главная биохимическая система к этому времени уже сложилась.
Тот факт, что она оставалась без изменений столь долго, определялся, без сомнения, взаимосвязанностью ее компонентов и сложностью. Это сложность того же типа, что и в инженерных разработках высшего уровня, когда многие тщательно отобранные компоненты настолько тесно взаимосвязаны, что ни один из них не может быть изменен. Такая «умная машина» могла быть лишь продуктом эволюции. В то же время далеко не ясно, сложился ли в результате эволюции и набор вошедших в ее состав компонентов. Итак, можно заключить, что единство биохимической организации сложилось не на начальных этапах эволюции, а гораздо позже.
ГЛИНА КРИСТАЛЛИЗУЕТСЯ из слабых растворов, которые образуются при просачивании воды сквозь выветренные породы. Слева кристаллы галойзитовой глины, которые растут в воде, просачивающейся сквозь трещины в граните (электронная микрофотография: увеличение х 3750). Справа кристаллы иллита, растущие в порах песчаника (увеличение х 16 000). Процессы такого рода могли играть главенствующую роль при возникновении жизни.
Второй постулат из вышеприведенной пары можно перефразировать так: «некоторые из важнейших биохимических компонентов живого получить очень легко — и точка». На доказательство и дополнение его новыми данными были нацелены эксперименты, выполненные вслед за экспериментами Миллера. Действительно, оказалось, что ряд веществ, к числу которых принадлежат некоторые простейшие аминокислоты, легко получить в самых разнообразных условиях, а не только сходных с существовавшими на ранних этапах эволюции Земли. Это «и точка» в постулате смазывает историческую перспективу, поскольку, приняв постулат, мы должны будем предполагать, что суть длительного эволюционного процесса заключалась в объединении компонентов, иные из которых было не слишком сложно соединить вместе и которые обладали достаточной стабильностью. Как и в случае первого постулата, все это мало говорит нам о событиях, происходивших у основания эволюционного древа.
Конечно, мы можем продолжать считать, что первые организмы были образованы из молекул тех же типов, что и современные, но такую точку зрения нужно рассматривать лишь как гипотезу, не имеющую подтверждений.
На самом деле имеется много оснований для сомнений. Прежде всего вспомним об исключительной сложности и взаимосвязанности основных биохимических процессов. Первые организмы не могли быть устроены таким образом. В них должны были действовать относительно простые «молекулярные машины», которые могли без особого труда вступать во взаимодействие и которые «работали» проще (выражаясь фигурально, это были копья, а не пулеметы). Здесь мы имеем дело с иным подходом к проблеме: он должен привести к мысли, что первые организмы имели другое строение, чем современные, и состояли из других веществ. Это справедливо и для вещей, сделанных человеком: для создания сложных и простых устройств нужны и разные материалы, и разные составные части. Не пытайтесь отыскать хоть одну костяшку от деревянных счетов в карманном компьютере (да и дерево вообще).
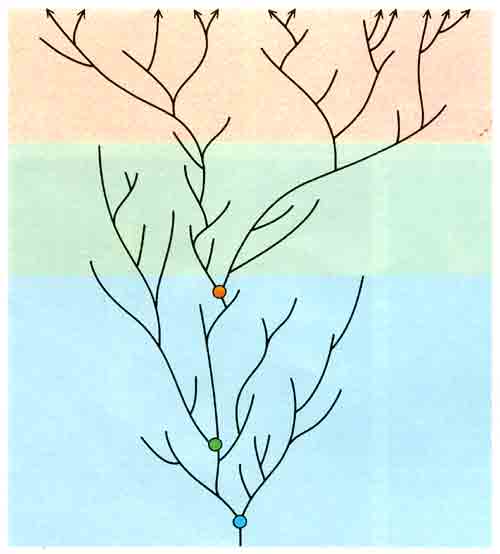
ГИПОТЕТИЧЕСКОЕ ЭВОЛЮЦИОННОЕ ДРЕВО: таким оно получилось в результате использования метода, согласно которому разветвление и вымирание отдельных ветвей происходят случайным образом. Оно очень схоже с другими древами такого рода тем, что все ныне существующие виды (показаны наверху стрелками) происходят из предковой точки ветвления, находящейся на некотором расстоянии от корня древа. (По мере роста древа такие точки ветвления могут переходить лишь на более высокий уровень.) Реально эволюция также идет путем ветвления и вымирания, поэтому неудивительно, что все организмы, существующие сегодня на Земле, вероятно, имеют относительно высоко развитого общего предка. Во всех ныне живущих организмах работает сложная молекулярная «машина», но из этого вовсе не следует, что такая же «машина» была характерна и для самых первых организмов.
Можно усомниться поэтому, что аминокислоты, которые замечательно подходят для построения катализаторов, нужных для определенных процессв, были столь же хороши для образования их в самом начале. Зададимся вопросом, действительно ли аминокислоты или другие важнейшие компоненты современного нам живого вещества вообще участвовали в «старте» жизни.
ОЧЕВИДНО, что современные живыe существа на всех уровнях их организации устроены очень сложно. Классическим примером сложного многокомпонентного устройства может служить глаз, который, для того чтобы вообше иметь практическую ценность, должен быть устроен именно так, как он устроен. «Как может такой орган возникнуть за счет малых эволюционных изменений?» - спрашивают антидарвинисты, думая, что попали в цель. Пусть они успокоятся: здесь нет ничего парадоксального. Высокий уровень организации может возникать за счет постепенных эволюционных изменений.
Представьте себе простои аналог «парадоксальной» структуры - каменную арку. Как можно построить арку постепенно, камень за камнем? Ответ: ее нужно собирать на опоре. А для начала нужно собрать эту опору, в которой нет ничего парадоксального, причем собрать по кусочкам - элемент за элементом.
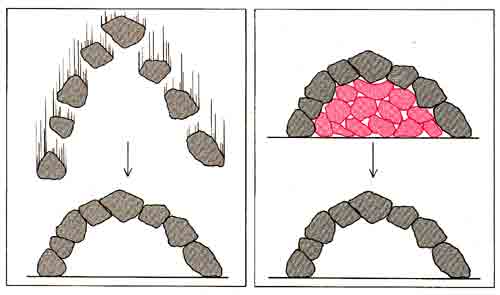
Я думаю, что именно таким путем и возникла известная нам, еще более удивительная, чем арка, биохимическая организация живого. Части ее, столь тесно пригнанные друг к другу сегодня, на первых порах опирались на что-то другое, устроенное более просто. Быть может, какие-то фрагменты древней опоры существуют и теперь, сама же опора разрушилась.
Что же дальше? Что мы можем сказать о том, чего уже нет?
Зададим иной вопрос: как узнать, каким оружием пользовались люди в древности, если не сохранилось никаких следов их деятельности? Вряд ли первым на ум придет пулемет, сделанный из камней и палок. Скорее вы попытаетесь представить наиболее простой путь, по которому могли пойти древние люди в создании оружия. При этом вы будете руководствоваться тем, что вы знаете о их потребностях, о технологии и доступных им материалах.
Рассуждая подобным образом, мы можем сказать о первых организмах следующее.
1. Они могли эволюционировать.
2. Они были просто организованы.
3. Они состояли из веществ геохимического происхождения.
Эти тезисы кажутся мне куда более правдоподобными, чем ранее рассмотренные, в них больше смысла и их стоит развить.
За тезис 1 можно не беспокоиться, так как я определяю организмы как системы, способные эволюционировать. Однако нужно быть очень внимательным в отношении смысла термина «эволюционировать»; это в конечном счете приводит к весьма точному описанию класса систем, к которым относятся самые первые организмы. Один организм эволюционировать не может: на это способны лишь организмы, некая их последовательность. Но и этого недостаточно. Эволюционировать может то, что объединяет организмы в ряд сменяющихся поколений, что передается от одного их поколения к другому. Этим является не вещество, а генетическая информация, не субстанция, а форма.
Очевидно, что генетическая информация должна храниться в каком-то материальном субстрате, в неких генах. И генетическая информация должна как-то проявляться (т.е. давать фенотип), что способствует ее сохранению и размножению. В этом, вероятно, принимают участие и другие вещества, но единственное, что сохраняется долго, — это сама информация. За время, необходимое для смены сотен поколений, каждый атом из того набора, который имелся у организма-основателя, будет заменен другим, исходное вещество исчезнет. Выживают только формы, измененные или неизмененные. Такой ход вещей, такой способ существования форм во времени путем многократного копирования и составляет сущность эволюции.
Есть и другие условия, определяющие саму возможность эволюции. В генетической информации должны происходить случайные изменения — мутации, и эти изменения должны наследоваться потомками и приводить к появлению измененных фенотипов; поэтому должен иметь место отбор измененной генетической информации. Таким путем за много поколений осуществляется изменение линий развития, и генетическая информация видоизменяется таким образом, что образуются фенотипы, которые особенно приспособлены к определенным условиям среды.
Конечно, мы сказали об эволюции далеко не все, что можно, но уже вправе сделать одно важное заключение: каков бы ни был состав первых, простых организмов, которые мы пытаемся себе представить, они должны были содержать некое подобие генов.
ЧТО ЖЕ еще, кроме генов, нужно было иметь этим первым организмам? Ответ на этот вопрос дал в 1926 г. Г. Мёллер, и звучал он так: «Ничего». Даже то немногое, что уже было известно и использовалось для объяснения свойств генов, оказалось в принципе достаточным для того, чтобы утверждать, что гены могут эволюционировать сами по себе. Мёллер пошел дальше. То, что первые организмы были просто генами, возможно не только теоретически — вероятно, они действительно представляли собой что-то в этом роде.
В первую очередь Мёллер подчеркивал, что иметь ген (или гены) было абсолютно необходимо. Предположим далее, что в первых организмах было и что-то еще. Для того чтобы эти организмы размножались, такие сопутствующие образования должны были либо синтезироваться заново, либо откуда-то поступать. Из чего следует, что для осуществления синтеза или приобретения требовалась дополнительная информация, которая должна была предсуществовать в генах. Конечно, лучше обходиться без «помощи» такого рода, а если уж прибегать к ней, то как можно реже.
Неоднократно высказывалось предположение, что в роли первичного вещества генов выступала РНК, хотя я считаю, что для этого она очень уж сложно устроена. И все же было показано, что молекулы РНК могут эволюционировать даже в пробирке. Важной особенностью РНК является то, что, хотя она, как и ДНК, несет генетическую информацию, способную к репликации, для того чтобы заключенная в РНК информация была эффективной, не обязателен процесс трансляции. Так же как и в случае белков, способ пространственного складывания цепей РНК может определяться содержащейся в ней наследственной информацией. Эти экспериментальные результаты очень интересны, но говорят ли они нам что-нибудь о ранних этапах эволюции? Фермент, который был использован в опытах с РНК, слишком сложно устроен , чтобы его можно было считать продуктом геохимических процессов, проходивших на ранних этапах эволюции Земли. Если бы даже для этой цели оказались пригодными более просто устроенные катализаторы, то на пути встала бы новая сложность: для репликации РНК нужны особые, макроэргические (богатые энергией) формы нуклеотидов.
В любом случае наш второй тезис побуждает мыслить в строго определенном направлении: ясно, что при попытке очертить особенности строения первых организмов вряд ли стоит использовать какие-то усеченные модели современной жизни. Поскольку первые организмы были просто устроены, они должны были быть и иначе устроены, и могли включать в свой состав совсем иные вещества. Возможно, в частности, что строение генетического материала, позволявшее обходиться без вспомогательных структур, отличалось от того его строения, которое стало возможным при возникновении таких сопутствующих структур в ходе эволюции.
Не так уж трудно представить себе эволюционный процесс, в результате которого первичный, геохимический генетический материал был постепенно замещен совсем другим материалом — органохимической природы. Я называю этот процесс генетическим захватом.
Если на ранних этапах эволюции центральной биохимической контролирующей машины действительно происходил генетический захват (или захваты), то вряд ли можно ожидать, что компоненты первичного генетического материала сохранились в современном молекулярном конструкторе. На первый взгляд это соображение подрывает гипотезу генетического захвата. Но у последней есть и достоинства: если следовать такой модели, открывается возможность использовать совершенно новые представления, предлагаемые химией. Наш третий тезис концентрирует внимание на мире минералов; при этом мы не оставляем в стороне основные соображения о том, каковы должны быть самые общие свойства генетического материала.
Вот что говорил Мёллер о природе генетического материала четверть века назад, еще до того, как стала известна роль ДНК: «В роли вещества генов может выступать любое соединение, которое в определенных условиях (в протоплазме или где-то еще) способно самовоспроизводиться с сохранением специфического состава и которое, кроме того, периодически изменяется — мутирует — и тем не менее сохраняет способность к самовоспроизведению во всем разнообразии своих форм».
Вывод о том, что в процессе репликации генов должны фигурировать какие-то матрицы, следует из этого высказывания со всей очевидностью. Трудно не увидеть в «специфическом составе» (генетической информации) некую специфическую пространственную организацию (паттерны), которая копируется за счет специфического расположения и связывания воедино контактирующих с нею мономеров. (Именно так обстоит дело при репликации ДНК и РНК.) Если матричный синтез представляется и не единственно возможным путем репликации сложных, мутабельных структур, то он во всяком случае принадлежит к числу простейших и наиболее прямых.
Теперь нам стоит поразмыслить о генетическом материале, составные части (мономеры) которого устроены проще, чем у ДНК. Нам нужно представить такой тип мономеров, которые могли бы образовываться на Земле с легкостью и постоянно в течение длительного времени. Специфических помощников — ферментов — тогда не было: компоненты первичного генетического материала должны были в той или иной мере обходиться самосборкой.
Теперь мы рассмотрим еще один тезис:
4. Гены должны включать большое число атомов.
Ген никогда не мог быть малочисленным объединением атомов, так как должен был содержать информацию в количестве выше среднего уровня, что позволяло ему эволюционировать. Более того, гены должны были представлять собой хорошо упорядоченные образования.
Не были ли первые вещества наследственности кристаллами? Ведь кристаллы — это наиболее часто встречающиеся образования, способные к самосборке. Аналогия между процессом кристаллизации и основными процессами жизнедеятельности проводилась неоднократно, но в конечном счете от нее отказывались как от слишком приблизительной. (Дж. Бернал пошел еще дальше: по его мнению, «кристаллизация - это смерть».)
На мои взгляд, возражения такого типа скрывают еще два ложных постулата:
3) структуры кристаллов слишком однообразны;
4) для живого лучше всего подходит углерод.
Первый из них не может рассматриваться всерьез, потому что корни его лежат в представлении о существовании идеальных кристаллов, которых на самом деле не бывает. Верно то, что у кристаллов есть основной тип строения, для которого характерна высокая периодичность, но в каждом реальном кристалле эта структура имеет дефекты. Даже сама конечность объекта (то, что он имеет форму и размер) - это уже «дефект», хотя, почти наверняка, найдутся и многие другие такие особенности. Некоторые структурные блоки могут отсутствовать или замещаться другими; образно говоря, большие или меньшие «куски обоев» могут быть смещены относительно друг друга в той или иной степени. Некоторые из таких нарушении могут быть весьма незначительными. Все это делает реальные кристаллы потенциально высоко информационноемкими.
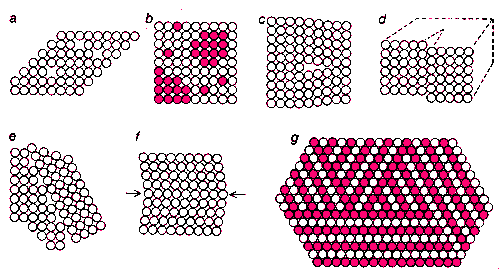
ДЕФЕКТЫ В КРИСТАЛЛАХ могут приводить к образованию множества стабильных альтернативных конфигураций, что является необходимым условием для хранения информации. На рисунке показаны наиболее обычные дефекты кристаллов: незанятые места в решетке (а), замещение отдельных элементов или доменов (Ь), краевые дислокации (с), вращательные дислокации (tf) и зернистые границы между решетками (е). В двойниковых кристаллах (f) по-разному ориентированные их части имеют общую плоскость взаимодействующих составляющих (стрелки). В некоторых кристаллах (g) большие домены имеют один и тот же состав, но различаются упаковкой составляющих их элементов.
Можно ли представить себе какие-либо структуры с дефектами, которые бы реплицировались в процессе роста кристалла? Ответ будет положительным. Это может происходить в кристаллах нескольких типов, обладающих подходящей комбинацией особенностей структуры, закономерностей роста и свойств спайности.
Остается разобраться с последним тезисом. Здесь мы опять-таки имеем дело не с истиной, а с предположением. Можно согласиться, что молекулы органических соединений — это наилучшие вещества для жизни. Но наилучшим, надо думать, является то, к чему эволюция пришла; начиналась же она, как мы вправе предположить, с наиболее доступного. Наиболее же доступная форма самосборки — это спонтанная кристаллизация простых, распространенных мономеров. Все эти рассуждения приводят к глине.
ВОКРУГ нас постоянно происходит процесс кристаллизации минералов глины из слабых растворов кремниевой кислоты и гидратированных ионов металлов, которые образуются при выветривании горных пород. В целом поверхность Земли можно уподобить гигантской фабрике по производству минералов глины.
Движущих сил здесь две. Во-первых, имеется геологический цикл, существующий за счет энергии радиоактивного распада, разогревающей недра. Этот цикл включает целый ряд процессов, в которых происходит захоронение осадочных пород, их преобразование при высоких температурах, погружение в более глубокие горизонты и наконец выход видоизмененных пород обратно на поверхность. В этих условиях они оказываются не полностью стабильными и с легкостью растворяются водой с образованием простейших составляющих — кремниевой кислоты и ионов металлов, которые могут кристаллизоваться, образуя совсем новые вещества — разнообразные минералы глины. В конце концов эти минералы в более или менее измененном состоянии вновь попадают в осадочные породы и опять подвергаются захоронению. За счет второго цикла обеспечивается поступление воды. Этот цикл зависит от Солнца: вода испаряется из морей, возникают облака, идут дожди, образуются грунтовые воды, ручьи и реки, а завершается все опять в морях.
Конечно, условия на Земле во время зарождения на ней жизни были иные. Из того, что мы знаем, впрочем, следует, что эти отличия не так уж велики. Метаморфизированные осадочные породы принадлежат к числу древнейших; это позволяет предположить, что климатический цикл действовал уже 3,8 млрд. лет назад. Вполне вероятно, что жизнь древнее таких пород. Возможно, она возникла в условиях, когда глины еще не могли формироваться, но для этой гипотезы нет особых оснований.
Не менее вероятно, что примитивные гены представляли собой микрокристаллические минералы, отличные от слоистых силикатов, т.е. «глин» в более широком понимании этого слова. Тем не менее я буду рассматривать именно обычные минералы глин.
ОДНО из следствий принятой мною линии рассуждений таково: простейший генетический материал (или что-то подобное ему) должен и сегодня образовываться на Земле. Как приступить к поиску этого материала? Из абстрактных описаний генетических кристаллов разных типов и из того, что мы знаем о минералах глин, мы можем составить представление о том, как они должны выглядеть, и заняться поисками реальных глин, которые соответствуют этим представлениям.
Рассмотрим вначале одномерный ген. Он должен быть обычен для глин, которые, по-разному сочетаясь, могут давать различные слоистые структуры. Возможна ситуация, когда слои одного типа наложены друг на друга различным образом или когда имеется последовательность слоев разных типов. Если вспомнить, что кристаллит глины, даже построенный из нескольких слоев, может быть весьма гибким и что одномерный кристаллический ген способен расти только в стороны, на ум легко придет некая структура, образованная определенным образом свернутыми (а может быть, и разветвленными) мембранами или лентами постоянной толщины. Существует множество глин с нерегулярной упаковкой слоев, которые весьма схожи с этим описанием.
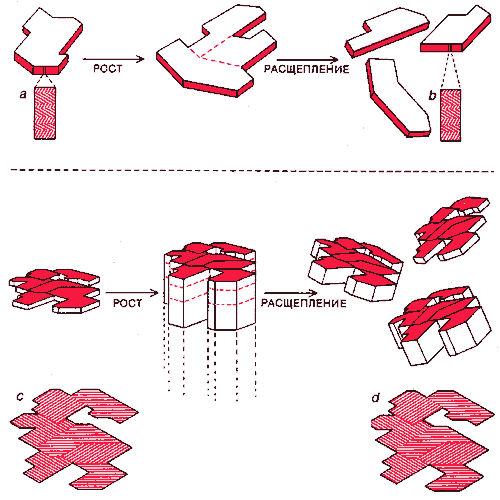
КРИСТАЛЛИЧЕСКИЕ ГЕНЫ должны были обладать правильной комбинацией структурных и ростовых признаков и признаков делимости. Информация могла храниться в одномерных или двумерных кристаллических генах. В случае одномерного гена (вверху) она содержалась в детальной структуре ряда взаимодействующих слоев (выделено цветом), которая сохранялась при репликации генов (а, b). Рост происходил только по окрашенным граням, а разделение шло только в параллельном им направлении. Физические свойства слоев, несущих информацию, могли быть различными (например, это могли быть по-разному уложенные кристаллические структуры); то же касается их химического состава. В двумерных кристаллических генах (внизу) информация могла храниться в форме специфической пространственной организации (в отношении и физических, и химических свойств) на грани кристалла (показано цветом). Эта организация сохранялась при репликации гена (с, d) за счет роста на окрашенной грани и при его расщеплении по плоскости, параллельной этой грани.
Конечно, во всем сказанном содержится лишь намек на истину. (Но как был бы интересен такой ген, способный неограниченно распространять содержащуюся в нем информацию без деления!) Рассмотрим теперь еще одну глину особого строения, которая может быть примером двумерного гена.
С. Бейли и К. Мэнсфилд, сотрудники Висконсинского университета в Мадисоне, провели рентгеноструктурный анализ крупных кристаллов вермиформного (червеобразного) каолинита и обнаружили в них интересные дефекты структуры. Отдельные слои каолинита представляют собой мозаично расположенные небольшие домены, составляющие в целом весьма прихотливый рисунок. В каждом домене все атомы алюминия имеют одну из трех возможных ориентации. В таких структурах может храниться очень много информации, и эта информация может быть реплицирована при условии, что во вновь образующемся слое ориентация атомов алюминия будет зависеть от их ориентации в том слое, на котором он формируется. В идеальных кристаллах каолинита ориентация атомов алюминия в слоях сохраняется, но в реальных кристаллах нередки «ошибки».
Сходный тип организации, видимо, присущ индивидуальным слоям типичных червеобразных кристаллов: для некоторых из них характерна сложная, но неизменная организация поперечного сечения. Как эти особенности строения, так и наличие глубоких борозд на поверхности говорят о доменной структуре (см. рисунок на с, 50).
Сказанное подтверждают результаты экспериментов А. Вепса из Мюнхенского университета, который исследовал рост кристаллов смектита. Вейс указывает, что новые слои, образующиеся в промежутках между предсуществовавшими слоями кристаллов, получают от них информацию: речь идет о распределении плотности отрицательных зарядов, появляющихся при замещении атомов на алюминий в кремний-кислородной сети.
Конечно, нужно проводить новые наблюдения и ставить опыты, чтобы обосновать правомерность главного вопроса: существуют ли минеральные, кристаллические гены? Сегодня на этот вопрос я могу ответить лишь так: «Быть может, существуют» — и перейти к другому вопросу: могли ли такие гены эволюционировать? На этот вопрос, как мне кажется, можно ответить так: «Да; маловероятно, что они не эволюционировали».
РАССМОТРИМ обычные микроусловия формирования глин — пористую структуру песчаника. Растворы, содержащие продукты выветривания, просачиваются сквозь него, и в порах происходит кристаллизация двумерно реплицирующихся глин. Процесс может одновременно происходить в ряде участков, и в каждом из них будут находиться миллионы кристаллов с определенными дефектами структуры. В каких-то из этих участков кристаллы могут иметь такие форму и размеры, что они будут близко прилегать друг к другу и образовывать водонепроницаемый заслон. Движение растворов в таких участках нарушится и кристаллы в них перестанут расти. В других участках небольшие, рыхло расположенные кристаллы не будут мешать протеканию растворов, но при дожде такие структуры будут легко вымываться, так что в этом тоже мало хорошего. Наконец, в участках третьего, типа кристаллы могут иметь такую форму, что они закрепятся в углублениях стенок пор; такие кристаллы и останутся на месте, не препятствуя протеканию питающих растворов. Возможны и участки, где реплицируются относительно длинные кристаллы, которые ввиду ограниченности пространства соединятся друг с другом и образуют довольно рыхлую структуру — это еще один путь сохранения постоянной локализации при поддержании потока питающих растворов.
Даже в таких относительно простых условиях могут проявляться весьма тонко действующие силы отбора — некие факторы, обусловливающие причины преимущественного развития определенных структур с дефектами (в данном случае имеются в виду главным образом их форма и размеры). В результате мутаций появятся варианты таких структур; по этой причине в разных частях растущих участков будут часто находиться кристаллы, несколько отличающиеся дефектами своей структуры. Это может привести к тому, что в одних местах рост пойдет быстрее, чем в других, одни части будут лучше переносить периодические неблагоприятные условия, чем другие.
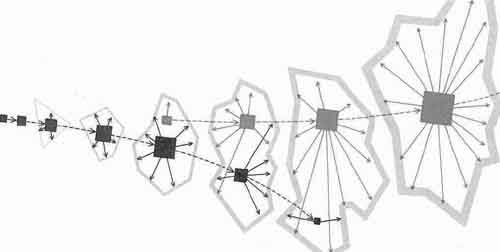
ГЕНЕТИЧЕСКИЙ ЗАХВАТ, каким он представляется автору, был важнейшим этапом ранней эволюции. Вначале существовали лишь «голые» гены, состоявшие из какого-то не известного нам первичного генетического вещества (черные квадратики слева), которые эволюционировали таким образом, что стали контролировать условия окружающей их среды путем управления образованием все более сложных, окружающих их фенотипов {голубые многоугольники). Возник новый тип генов {красные квадратики), которые могли «работать» только в границах довольно сложно устроенного фенотипа, но были более «работоспособными», чем исходные гены. Новые гены постепенно захватывали контрольные функции при формировании фенотипов, так что в конечном счете начальная форма генов была ими полностью вытеснена.
Давайте теперь рассмотрим несколько более сложные условия. Пусть почвенные растворы из песчаника попадают в быстро текущий поток, который в силу повышенной кислотности и малой концентрации веществ в нем не обеспечивает условий для синтеза глин, но содержит один из компонентов (нужных для образования «генетической» глины), которого недостает в песчанике. С позиции химии идеальным местом для образования такой глины будет в этом случае поверхность раздела между двумя разными средами. С физической точки зрения условия для этого отнюдь не идеальны, так как эта поверхность невелика и подвержена изменениям. Образующиеся кристаллы всегда находятся под угрозой вымы-вания или растворения либо того и другого вместе. Тем не менее поверхность раздела может быть стабилизирована и расширена значительными группами тесно взаимодействующих (реплицирующихся) кристаллов, прикрепленных к песчанику; получится весьма клейкая масса. Мутации, изменяющие форму образующих ее переплетающихся кристаллов, будут изменять ее пористость, что в свою очередь приведет к изменению в ней градиентов кислотности, а также градиентов концентрации различных ионов. Подбор условий, подходящих для синтеза глин в таких массах, может происходить путем естественного отбора. Наибольшие шансы на успех будет иметь такой «комок генов», который окажется и хорошей поддерживающей средой, и подходящим местом для образования глины. Считается, что длинная шея у жирафа возникла под действием естественного отбора; в результате животные этого вида могут поедать листья с макушек деревьев без особых усилий. Можно думать, что и разные формы реплицирующихся глинистых частиц могут в определенных условиях также «приспосабливать» свой размер и форму. Забудем на время о сложностях физиологии. В логике событий здесь действительно есть много общего. Реплицирующиеся, мутирующие структуры оптимизируются одним и тем же путем — в ответ на меняющиеся условия. Они просто не могут вести себя иначе.
ДО СИХ ПОР мы рассматривали самые первые организмы как состоящие из одних только генов. Такие организмы действительно могли существовать, поскольку даже сам набор генов может обладать фенотипом. Иными словами, он характеризуется неким комплексом физико-химических свойств, определяемых имеющейся в генах информацией, от которых зависит его эволюционная судьба. В реальных условиях исходный набор «глиняных» генов постоянно «загрязнялся» бы другими глинами, образовавшимися в условиях, которые возникли под влиянием этих генов. Совместная кристаллизация могла иногда приводить к улучшению свойств агрегатов, например их пористости или способности удерживаться в месте роста. Одновременно могли происходить мутации генов, попадавшие под действие отбора, который способствовал образованию «вторичных» глин. Нетрудно представить себе, каким образом дефектная структура в генетическом кристалле могла самым прямым способом осуществлять контроль над ростом аналогичных образований: это могло происходить за счет эпитаксии, т.е. под влиянием специфических дефектов предсуществовавших структур на кристаллизацию вторичных глин на поверхности генетических глин.
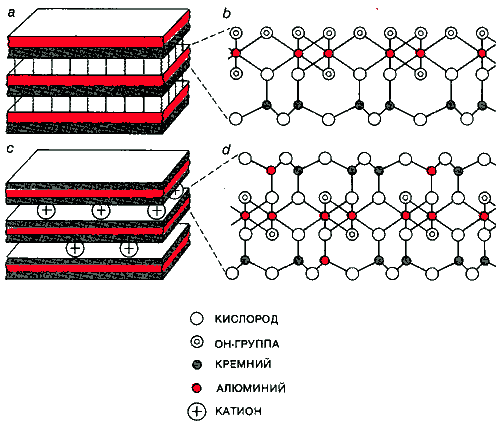
БОЛЬШИНСТВО ГЛИН образовано стопкой слоев. В каолините (а) асимметричные слои скреплены водородными связями. Каждый слой образован сеткой из атомов алюминия и гидроксильных групп, слитой воедино с сетью из атомов кремния и кислорода (b). В других глинах слои симметричны. В них кремний-кислородная сеть с обеих сторон слита с металл-гидроксильной сетью. Эти слои заряжены отрицательно и взаимодействуют при участии положительно заряженных ионов (с). В иллитах (d) большая часть отрицательных зарядов появляется в результате замещения атомов кремния атомами алюминия.
Рассмотрев первые организмы, пойдем теперь дальше. Я попробую в нескольких словах описать взаимосвязи между нами и первыми организмами. Для этого нам не обойтись без молекул органических соединении, а также без вопросов «почему?» и «как?».
Почему вообще на сцену вышли молекулы органических соединений? По ряду причин. Некоторые из малых органических молекул (например, аминокислоты или ди- и трикарбоновые кислоты) могли способствовать солюбилизации ионов металлов, например алюминия. Таким образом, они выступали в роли катализаторов при синтезе глин. Молекулы других типов (например, гетероциклические основания и полифосфаты) обладают повышенным сродством к глинам, что часто приводит к изменению физических свойств геля глины. Органические молекулы способны также сильно влиять на форму и размер неорганических кристаллов, подавляя рост некоторых их граней. Это могло иметь особо большое значение для контролируемой репликации кристаллических генов. Кроме того, влияние на структуры могли иметь и полимерные органические соединения. Они могли удерживать частицы глины рядом друг с другом.
Я думаю, что предшественники РНК, появившиеся в достаточно развитых глиняных организмах, в первую очередь играли структурную роль. (На самом деле РНК и сегодня иногда используется приблизительно для тех же пелей.) РНК-подобный полимер с отрицательно заряженным остовом молекулы должен стремиться связываться с краями частиц глины (они чаще всего заряжены положительно). А гетероциклические основания (молекулы типа аденина) имеют тенденцию проникать между слоями глины. Можно представить себе некий РНК-подобный полимер, который возник специально для взаимодействия с глинами (возможно, даже для «чтения» информации, экспонированной на краях одномерных «глиняных» генов).
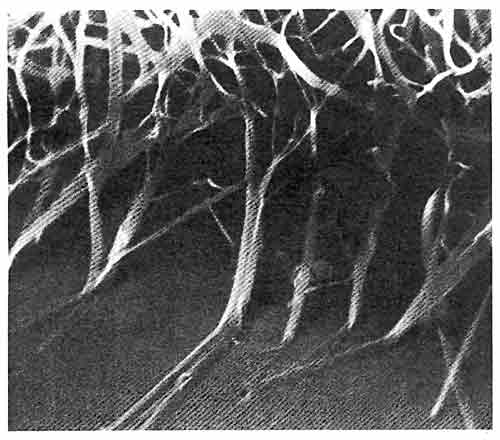
ДЛИННЫЕ КРИСТАЛЛЫ ИЛЛИТА, прикрепленные к зерну песчаника. (Увеличение х 10 000; микрофотография сделана при помощи сканирующего электронного микроскопа.) Такая глина — подходящий кандидат на роль одномерного генетического кристалла, но, поскольку она образована небольшим числом слоев, ее информационная емкость невелика.

ВЕРМИФОРМНЫЙ (ЧЕРВЕОБРАЗНЫЙ) КАОЛИНИТ (увеличение х 1350), образовавшийся путем выветривания. Такая глина — возможный кандидат на роль двумерного генетического кристалла.
Генетический захват, который привел к становлению существующих сегодня механизмов биохимического контроля, мог начаться, согласно этой гипотезе, с того момента, когда РНК стала реплицирующейся молекулой. То был новый тип «сотрудничающего» генетического материала, роль которого вначале была невелика. Для того чтобы отойти от основы — глиняного каркаса, — требовалась длительная эволюция. Возможно, это произошло уже после того, как возникла сложная система механизмов белкового синтеза. Об эволюции такой системы можно рассуждать, поскольку она происходила в пределах образовавшегося организма. Сначала она могла эволюционировать как побочная система, но постепенно становилась все более полезной и сложной, а когда каркас исчез, стала совершенно необходимой.
Что было причиной перехода эволюции с неорганического кристаллического уровня на органический молекулярный? Дело, видимо, в том, что органические вещества могут иметь более тонкую структуру (конечно, если есть соответствующие механизмы для ее создания), что позволяет осуществлять более сложный контроль над процессами.
В какой последовательности вступали в действие органические молекулы? Мне кажется, что все началось с фотосинтеза, в котором использовался углекислый газ атмосферы и который на первых порах приводил к образованию молекул типа муравьиной кислоты.
Однако значительно интереснее другой вопрос — о том, каким образом происходило объединение более сложно устроенных молекул до появления ферментов. Как происходило образование нуклеотидов? Для этого должны были сложиться необходимые для их формирования системы, включающие многие химические реакции и другие процессы, например процессы очистки; причем все должно было осуществляться в строго заданной последовательности. Такая система не могла быть результатом случайных процессов: она должна была быть организована. У достаточно развитых организмов, «сделанных» из глины, в роли организующего начала мог выступать естественный отбор. С какими же физическими объектами имел он дело до того, как к работе приступили молекулы белков? Я думаю, что до появления ферментов в организмах действовал какой-то более «старомодный» аппарат, в чем-то схожий с теми, которые имеются в лабораториях химиков-органиков или химических заводов. В них наряду с колбами, пробирками, насосами, ионообменными и адсорбционными колонками широко применяются и весьма малоспецифичные катализаторы.
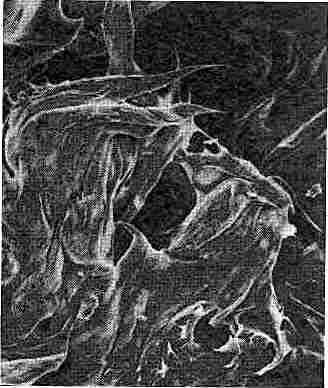

РАЗНООБРАЗИЕ ФОРМ минералов глины — один из фактов, говорящих о возможной роли глин в построении примитивных организмов. Кожистый галлойзит показан на этом рисунке с разным увеличением: он образован массой нитей, часть которых представляет собой полые трубочки. (Микрофотографии сделаны при помощи электронного микроскопа.) Очень маленький отросток едва заметен при увеличении в 130 раз (слева). При увеличении в 1000 раз на нем выявляется петля (е середине). При увеличении в 26 000 раз видно, что он представляет собой полую трубочку (справа).
С учетом сказанного мы переходим к заключительной части представлений о минералах глин как основных веществах, из которых сформировались первые организмы. Коль скоро нам нужны катализаторы (довольно неспецифические), мы легко обнаружим их среди минералов глин. Точнее, если нам нужен достаточно простой аппарат типа тех, о которых только что шла речь, то существует множество обычных глин, которые могли бы быть использованы для его построения. Как его части объединились и какие силы отбора действовали при этом — неясно.
Скептики могут задать три вопроса:
I. Почему кристаллические гены не являются обычным компонентом окружающей нас природы, если они на самом деле состоят из простых веществ и если их эволюция — это естественный процесс?
Я мог бы предложить четыре ответа на этот вопрос: 1) быть может, кристаллических генов вообще не существует; 2) генетические минеральные вещества встречаются весьма редко; 3) подходящие условия для репликации минеральных генов складываются нечасто; 4) минеральные гены, прошедшие определенный путь в эволюции, встречаются часто, но мы не умеем их различать. Оставляю выбор ответа читателю.
В любом случае я не могу представить себе, что современные организмы на основе глин могут вновь достигнуть такой фазы развития, что станут способны использовать органические молекулы. Слишком большую конкуренцию составляют им ДНК-содержашие микроорганизмы. То же самое можно сказать и о предковых формах: сегодня их также, наверное, не существует.
II. Имеем ли мы право приписывать маленьким кристаллам глин свойства живого?
Первые организмы были, скорее всего, весьма непредставительными и, я бы сказал, неживыми. Организм нужен как предпосылка эволюции, но «жизнь» — это что-то совсем иное, нечто необычное, некая на вид целенаправленная сложность, которая могла. сформироваться в результате эволюции. Но на более поздних стадиях первичные организмы, как мне кажется, должны быть уже живыми с любой точки зрения.
III. Какие эксперименты нужно провести?
Поверхность раздела между глиной и молекулами органических соединений в настоящее время интенсивно изучается. Эта работа в основном ведется в Эймсском центре НАСА, расположенном близ Маунтин-Вью в Калифорнии. Дж. Лоулесс и его коллеги выяснили, каким образом ионы металлов, таких, как медь и цинк, могут способствовать связыванию нуклеотидов с глинами. Они также обнаружили, что ионы глин обладают селективным каталитическим влиянием на аминокислоты. Работая в этом же центре, М. Мортланд из Университета шт. Мичиган обнаружил, что кофермент пиридоксальфосфат может функционировать как фермент, если он находится в комплексе с медьсодержащими монтмориллонитовыми глинами. Н. Лахав из Еврейского университета вместе с Д. Уайтом из Университета в Санта-Клара и Ш. Чангом из Эймсского центра выяснили, каким образом глины, подвергающиеся периодическому смачиванию и высушиванию, могут способствовать соединению молекул аминокислоты глицина. В этом циклическом процессе происходит перенос энергии от окружающей среды к органическим молекулам.
Механизм преобразования энергии должен был быть необходимой составной частью организмов, за исключением самых примитивных. Л. Койн из Калифорнийского университета в Сан-Хосе нашла, что для этой цели вполне могли служить каолинитовые глины. Они могут собирать энергию из окружающей среды, которая выделяется при радиоактивном распаде, сохранять ее и высвобождать в тех случаях, когда структура глины нарушается определенным образом, например при ее смачивании или высушивании.
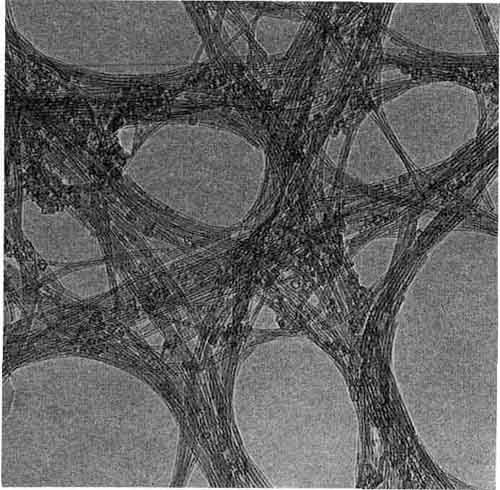
ИМОГОЛИТ И АЛЛОФАН (увеличение х 500 000; снимок сделан при помощи электронного микроскопа). Длинные тонкие образования - это монолитные трубочки из имоголита, они усеяны полыми «стручками» аллофана.
Пытаясь составить более определенное представление о взаимодействиях на границе раздела глина — органические молекулы, Лоулесс, Чанг и их сотрудники исследовали углеродсодержащие метеориты, столь же древние, как и сама Солнечная система. Они хотели найти ответ на вопрос: какова была органическая химия веществ на ранних этапах эволюции Земли? Интересно, что в таких метеоритах одновременно присутствуют и глины, и органические вещества. Изучение поверхности Марса также поможет нам понять, каковы были условия на начальных стадиях эволюции Земли. Э. Бейнин из Еврейского университета предлагает точку зрения, согласно которой на поверхности Марса встречаются главным образом богатые железом монтмориллонитовые глины. В этой связи объяснимы результаты опытов, выполненных посадочным аппаратом «Викинг», в которых под влиянием ультрафиолета происходило превращение углекислого газа и окиси углерода в молекулы органических соединений (среди которых большую часть составляла, видимо, муравьиная кислота). Дж. Хаббард из Технологического института в Джорджии поставил опыты, которые показали, что железосодержащие минералы, включая глины, дают тот же эффект.
Самая сложная и важная задача исследований сегодня — обнаружить кристаллические гены, и не одного типа, а многих типов, причем не только минеральные. Представьте себе эксперименты с кристаллами, способными к эволюции. Мы могли бы создавать разные формы давления отбора, и смотреть, как они ведут себя в таких условиях. Вне всякого сомнения, это было бы увлекательным занятием независимо от того, из чего состояли бы кристаллы. Мы очень скоро смогли бы решить, возможно ли существование реплицирующихся систем на минеральной основе, хотя в результате, быть может, мы бы и утратили интерес к самым отдаленным нашим предкам, ведь у нас в руках оказались бы совсем иные первичные организмы — первые организмы нашего собственного изобретения.
(Последние исправления - 7.3.2002)